Разведение и содержание раков
РАЗВЕДЕНИЕ И СОДЕРЖАНИЕ РАКОВ
Речные раки — очень ценные беспозвоночные животные, постоянно пользующиеся большим спросом во всех уголках земного шара. Но, к сожалению, природные популяции их ежегодно уменьшаются, чему способствуют браконьерство, периодически возникающие различные эпизоотии и другие причины. Установлено, что природные запасы раков достигают максимума каждые 7 лет, после чего начинают снижаться до минимума.
В последние годы большое внимание уделяется разведению раков в искусственных водоемах. Лидирует здесь Турция, куда в свое время были завезены раки из водоемов Украины. На внешний рынок она ежегодно поставляет до 7 тыс. тонн товарных раков; несколько меньше — Испания (3,5 тыс. тонн) и Китай (1 тыс. тонн).
В приусадебных и дачных прудах можно с успехом культивировать раков таких быстрорастущих видов, как широкопалый (Astacus astacus) и длиннопалый (Pontostacus lepto-dactylus).
Раки могут обитать в любых пресных водоемах — реках, озерах, лиманах, водохранилищах, прудах и т. п. Они любят чистые, не загрязненные химическими, бытовыми или промышленными отходами природные водоемы (не зря их считают индикаторами чистой воды), глубина которых обычно 2-5 м, но в отдельных случаях может доходить до 8-15 м. Идеальная среда для обитания раков — береговая линия водоема с затоками, где хорошо развивается водная растительность (элодея, хара, роголистник и др.), покрывающая (только не полностью) дно водоема. При поедании водорослей в организме животных ускоряется кальциевый обмен, а это способствует затвердению панциря после линьки. Раки предпочитают достаточно плотное дно (песчаное, глинистое) с присутствием известковых пород, камней, а также водоемы с нормативной или повышенной минерализацией воды. Грунт берега и дно водоема должны быть такими, чтобы ракам удобно было строить норы, хотя они могут обитать и под камнями, корнями, пнями.
Чаще всего раки строят норы на отвесных тенистых берегах, где мало солнца. Обычные размеры нор: длина — 7-36 см, ширина — 4-18 см, высота — 2-16 см. Зимой они расположены на большей глубине, а летом — на меньшей. Роют раки свои норы с помощью ног и хвоста, опираясь на передние клешни. Кстати, хвост им нужен не только для рытья нор, но и для плавания. Плавают раки, как известно, задом наперед и при этом бьют хвостом по воде. По дну они передвигаются медленно и часто также задом наперед. Случается, что раки покидают водоемы с загрязненной водой, передвигаясь по суше. В воде с кислой реакцией они, как правило, не живут. Оптимальное количество растворенного в воде кислорода для речных раков — 6-7 мг/л, однако допустимо кратковременное снижение его уровня до 2-3 мг/л.
Обычные размеры нор: длина — 7-36 см, ширина — 4-18 см, высота — 2-16 см. Зимой они расположены на большей глубине, а летом — на меньшей. Роют раки свои норы с помощью ног и хвоста, опираясь на передние клешни. Кстати, хвост им нужен не только для рытья нор, но и для плавания. Плавают раки, как известно, задом наперед и при этом бьют хвостом по воде. По дну они передвигаются медленно и часто также задом наперед. Случается, что раки покидают водоемы с загрязненной водой, передвигаясь по суше. В воде с кислой реакцией они, как правило, не живут. Оптимальное количество растворенного в воде кислорода для речных раков — 6-7 мг/л, однако допустимо кратковременное снижение его уровня до 2-3 мг/л.
Обычно раки ведут ночной образ жизни, но если они почуют добычу, то будут стремиться к ней, несмотря на время суток. Любопытен и такой факт: самки всегда сидят в норах поодиночке, а самцы во время зимовки нередко собираются группами и зарываются в ил.
Раки — раздельнополые животные. Самцы длиннопалых раков, например, достигают половозрелости на третий год жизни при длине тела 8 см, а самки — только на четвертый год при длине тела 7 см. Как правило, раки-самцы в 2-3 раза крупнее самок. Спаривание происходит либо осенью (октябрь—ноябрь), либо в конце зимы — начале весны (февраль—март) при температуре воды 10°С. Продолжительность спаривания — 2-3 недели, оплодотворение внешнее. Самцы приклеивают свои сперматофоры на нижней стороне головогруди самки в виде белого пятна. При позднем спаривании и низких температурах оплодотворение происходит за несколько дней.
Как правило, раки-самцы в 2-3 раза крупнее самок. Спаривание происходит либо осенью (октябрь—ноябрь), либо в конце зимы — начале весны (февраль—март) при температуре воды 10°С. Продолжительность спаривания — 2-3 недели, оплодотворение внешнее. Самцы приклеивают свои сперматофоры на нижней стороне головогруди самки в виде белого пятна. При позднем спаривании и низких температурах оплодотворение происходит за несколько дней.
Икру самки откладывают ночью, в тишине, в течение 2-3 часов. Подгибая брюшко к головогруди, они образуют камеру, в которую выпускают специальное вещество, растворяющее прикрепленные там сперматофоры со сперматозоидами. Икринки, выдавленные из яйцевода, проходят через семенной раствор, оплодотворяются и прикрепляются к брюшным ножкам или панцирю. Плодовитость самок зависит от их размера, физиологического состояния, времени года и других факторов. На ножках у них может находиться 110-480 икринок. Благодаря движениям ходильных и брюшных ножек икринки постоянно омываются свежей водой.
Маленькие рачки (личинки) выклевываются из икры чаще всего в третьей декаде мая и первой декаде июля при температуре 21-24 °С. Первые 2-3 дня они висят на так называемых гиалиновых нитях, затем нити обрываются, а рачки с помощью маленьких загнутых назад крючков на клешнях прикрепляются к оболочке яйца. В таком состоянии они находятся 5-8 дней, питаясь только за счет запасов желтка, который расположен под спинным панцирем головогруди малышей. До десятого дня происходит первая линька, после которой личинки становятся похожими на взрослых раков. Масса их составляет 21-30 мг, длина тела — 1,1-1,2 см. Питаются рачки самостоятельно, но при неблагоприятных условиях прячутся под брюшко матери.
На тринадцатый-двадцатый день происходит вторая линька, после которой личинки становятся полностью самостоятельными. В период линьки рачки находятся в укрытии. За это время они подрастают, выпрямляют ножки и начинают двигать клешнями, усами, глазами. Между головогрудным щитом и брюшком возникает щель, из которой рачки высовывают свое мягкое тело. Иногда клешни и ножки отрываются, но затем они восстанавливаются. Чаще всего сроки линьки приходятся на май — август. Линька у раков проходит многократно: на первом году жизни — 8 раз, на втором — 4-5 и на третьем — 3-4 раза. Первые 1-1,5 месяца жизни для маленьких рачков — очень опасный период: они особенно подвержены болезням, их могут съесть рыбы, ондатры, водоплавающая птица.
В период линьки рачки находятся в укрытии. За это время они подрастают, выпрямляют ножки и начинают двигать клешнями, усами, глазами. Между головогрудным щитом и брюшком возникает щель, из которой рачки высовывают свое мягкое тело. Иногда клешни и ножки отрываются, но затем они восстанавливаются. Чаще всего сроки линьки приходятся на май — август. Линька у раков проходит многократно: на первом году жизни — 8 раз, на втором — 4-5 и на третьем — 3-4 раза. Первые 1-1,5 месяца жизни для маленьких рачков — очень опасный период: они особенно подвержены болезням, их могут съесть рыбы, ондатры, водоплавающая птица.
Еще одна любопытная деталь из жизни раков. В поисках пищи они мигрируют, питаясь в основном беспозвоночными (червями, моллюсками, насекомыми и их личинками, мелкими ракообразными и др.), а также богатой кальцием водной растительностью, мелкой рыбешкой. Очень интересно наблюдать за процессом поедания раками пищи. Если добыча находится близко от норы, то они относят ее в свое укрытие, если далеко — рак съедает добычу на месте, прячась в любое укрытие.
Оптимальная температура окружающей среды для полноценного питания взрослых раков — 17-21°С, личинок — 18-23 °С. Несмотря на заботливый уход самок за своим потомством, раки могут поедать друг друга. Это явление обусловлено групповым образом жизни животных, частыми линьками, неодинаковым ростом. Крупные особи поедают более мелких.
Раки, как и все животные, болеют. Часто их поражают чума и ржавопятнистая болезнь. На панцире животных могут оседать различные моллюски (особенно дрейсены, нитчатые водоросли), но при линьке раки их сбрасывают.
Разведением речных раков занимаются во многих странах мира. Кроме Турции, Испании и Китая это Россия, США, Германия и др. В каждой стране разработана своя технология производства раков, но основные требования такие: наличие водоемов с малозаиленным глинистым дном и водой, обогащенной кислородом, поддержание постоянного летнего температурного и гидрохимического режима. Для разведения раков можно использовать специализированные, фермерские, а также небольшие частные и приусадебные участки, на которых имеются водоемы.
Существует два типа хозяйств по разведению раков — прудовой и заводской. Экономически наиболее выгодным считается первый, поскольку разведение раков — процесс довольно трудоемкий. Начинающим раководам целесообразно для начала выращивать сеголеток, реализация которых при постоянном рынке сбыта может дать значительную прибыль.
При разведении раков важное значение имеет заготовка самок с живой икрой на плеоподах (ножках) и их транспортировка в рачьи хозяйства. Чтобы вырастить 1 тонну раков, необходимо заготовить 500-600 самок; их отлавливают в природных водоемах. В хозяйстве (маленькие пруды, бассейны или специальные аппараты) проводят доинкубацию зародышей, находящихся на плеоподах. При этом очень важно создать хороший водообмен и аэрацию воды.
При разведении раков необходим постоянно наблюдать за качеством воды, контролировать количество растворенного в ней кислорода (не менее 5-7 мг/л) и водорода (7-9 мг/л). Водообмен должен составлять примерно 50 л/мин на 1000 кв.
Производителей раков помещают в пруды (площадь — около 0,1 га, глубина — 1-1,5 м, плотность посадки — 1-5 шт. на 1 кв. м). При температуре воды выше 7 °С ракам скармливают свежий или вареный корм (мясо, отходы боен, овощи, моллюски и т. п.). Средняя суточная норма должна составлять 2% массы тела рака. Корм размещают на деревянных лотках (40×40 см). При прудовом способе разведения раков- личинки первой стадии выходят в мае — июне. После второй линьки молодь (животные, еще не достигшие величины взрослых раков) отлавливают и пересаживают в маточный пруд, а маленьких рачков доращивают до сеголеток массой 7-10 г. Их можно доращивать в этом же пруду либо пересадить в другой, в котором условия удовлетворяют требованиям зимовки. Раков-годовиков отлавливают и пересаживают в нагульные пруды, где плотность посадки меньше, чем в предыдущем водоеме.
Содержать и разводить раков можно и в домашних условиях (аквариумах, ваннах), однако в этом случае получают лишь небольшое количество личинок (рачков). Много хлопот доставляют их кормление и сохранение. Таким образом, вырастить раков до товарной массы в домашних условиях очень сложно.
Чтобы получить 3-4 ц/га товарной продукции раков, необходимо иметь не меньше 3-4 прудов, подготовленных надлежащим образом.
Речные раки, как известно, в загрязненной воде не живут, поэтому мясо у них более чистое, чем у других водных животных. Да и сами они в пруду выполняют функцию санитаров, очищая водоемы от органических остатков.
В приусадебном пруду рекомендуется разводить широкопалых и длиннопалых раков. У широкопалого рака клешни широкие, панцирь гладкий, длина тела около 15см. Длинно-палый рак имеет узкие и длинные клешни и шероховатый панцирь; длина тела достигает 20 см, вес самца — более 300 г.
Зимой раки уходят на глубину и зарываются в ил. Здесь им комфортно, и пищи хватает. В зимний период, как известно, кислорода в воде недостаточно, некоторые рыбы задыхаются, падают на дно и становятся добычей раков, которые даже в условиях пониженной температуры не прекращают активно питаться.
Отлавливают раков специальными удочками, рачевнями и мережками, начиная с середины лета и до поздней осени. Хороший улов бывает в темных водах в вечерние часы, в прозрачных — с наступлением сумерек и до полуночи. Наилучшие уловы бывают в темные теплые ночи и в дождливую погоду.
Для того чтобы пойманные раки лучше сохранялись и не нападали друг на друга, их надо подкармливать крапивой, ольховыми листьями, картофелем и другой растительностью Свежую рыбу давать не рекомендуется, так как раки при этом устраивают потасовки, во время которых теряют клешни и ноги, а значит, товарный вид.
Тест по биологии Класс ракообразные 7 класс
Тест по биологии Класс ракообразные 7 класс с ответами. Тест включает 2 варианта, в каждом по 7 заданий.
Тест включает 2 варианта, в каждом по 7 заданий.
Вариант 1
A1. Кто не относится к ракообразным?
1) креветка
2) мокрица
3) щитень
4) перловица
А2. Что представляет собой выделительная система ракообразных?
1) почки
2) зеленые железы
3) трубочки
4) канальцы
А3. Из какого органического вещества состоит покров ракообразных?
1) из хитина
2) из кальция
3) из скорлупы
4) из мы щелока
А4. Как называются органы равновесия у ракообразных?
1) клешни
2) статоцисты
3) усики
4) подошва
B1. Что циркулирует в кровеносной системе ракообразных?
В2. С помощью чего измельчается пища у ракообразных в желудке?
C1. Как передвигается рак?
Вариант 2
A1. Кто не относится к ракообразным?
1) краб
2) скорпион
3) креветка
4) бокоплав
А2. Что представляет собой центральная нервная система ракообразных?
Что представляет собой центральная нервная система ракообразных?
1) нервные отростки
2) мозг
3) окологлоточное нервное кольцо, брюшная нервная цепочка
4) щупальца
А3. Чем дышат ракообразные?
1) легкими
2) жабрами
3) дыхательными трубочками
4) ртом
А4. Какой процесс характерен для ракообразных?
1) линька
2) спячка
3) паразитизм
4) эхолокация
В1. Как называются мелкие глазки, из которых составлен глаз ракообразных?
В2. Какова выделительная система ракообразных?
C1. Как от времени года зависит размножение раков?
Ответы на тест по биологии Класс ракообразные 7 класс
Вариант 1
А1-4
А2-2
А3-1
А4-2
В1. Гемолимфа
В2. С помощью жерновков
Вариант 2
А1-2
А2-3
А3-2
А4-1
В1. Фасетки
В2. Пара зеленых желез
способы разведения речных обитателей, как особи спариваются и откладывают икру
Раки считаются интересными и необыкновенными существами. Люди стараются разводить их в домашних условиях по разным причинам. Одни — чтобы разнообразить домашнюю фауну необычными питомцами, другие — для украшения своего меню, так как их мясо считается полезным деликатесом. Размножаться раки начинают с приходом весны. Их разведение в домашних условиях не отнимает много сил и не требует крупных финансовых вложений, но занимает много времени.
Люди стараются разводить их в домашних условиях по разным причинам. Одни — чтобы разнообразить домашнюю фауну необычными питомцами, другие — для украшения своего меню, так как их мясо считается полезным деликатесом. Размножаться раки начинают с приходом весны. Их разведение в домашних условиях не отнимает много сил и не требует крупных финансовых вложений, но занимает много времени.
Выведение потомства
Плодотворность зависит от количества особей разных полов. Самцы и самки имеют некоторые различия, по которым без особого труда можно поддерживать их оптимальное соотношение.
Самцы по своим габаритам крупнее женских особей в одинаковой возрастной категории. Отличаются они и расположением половых отверстий: у самок они находятся у третьей пары лап, а у представителей противоположного пола — возле пятой пары. На хвосте сразу за лапками у речных броненосцев находятся брюшные ножки. У самцов первые две пары хорошо развиты, а у самок они являются лишь мягким придатком или их нет вовсе.
При спаривании рак обхватывает самку ногами и крепко прижимается к внутренней её стороне. Происходит переливание спермы в самку и оплодотворение икры. Самка при спаривании активно отбивается от захвата самца. После оплодотворения она теряет силы из-за своего активного сопротивления. Потрепанная, она прячется в норе, выползая оттуда в поисках еды в дневное время.
Самец, наоборот, сохраняет активность и способен оплодотворить ещё нескольких самок. После нескольких спариваний он измождён и голоден. Бывает такое, что после последнего соития он может съесть свою избранницу.
Чтобы избежать подобных последствий, в водоёме нужно регулировать численность питомцев — она должна соответствовать пропорции 1:3.
Рост личинок
Интересно наблюдать за тем, как рождаются раки. Оплодотворённая самка через 18−20 дней начинает откладывать икру, которая приклеивается к плодоножкам на хвосте и держится там до появления личинок. Этот период для самки является самым тяжёлым. Она постоянно гоняет воду, насыщенную кислородом, быстро разгибая и подгибая хвост, чтобы промыть икру.
Когда самка долгое время находится в норе, вода там застаивается, что может привести к гибели икринок. А также их могут повредить мелкие насекомые, например водяные скорпионы, жуки-гладыши.
А также их могут повредить мелкие насекомые, например водяные скорпионы, жуки-гладыши.
Даже при постоянном промывании икры от плесени, грязи и водорослей некоторые икринки отрываются, падают на дно и пропадают. Из всего отложенного потомства появляются на свет около шестидесяти личинок, длина которых равна 1,5 мм.
Из всего отложенного потомства появляются на свет около шестидесяти личинокСначала они держатся на плесу у самки. Через две недели начинают плавать около неё, а при любой опасности мгновенно прячутся под её хвост. Набрав немного сил, малыши покидают самку и начинают самостоятельную жизнь.
Личинки растут медленно. В три месяца их длина составляет 3 см, а внешний вид начинает соответствовать взрослым особям. Через один год их длина равна 4 см, через два года они вырастают до 6 см.
Чтобы выйти из икринки, рачок ножками и брюшком разрывает оболочку и зависает на нити. Через три дня нить обрывается, и личинка острыми клешнями схватывается за оболочку икры или водоросль. Так маленький представитель ракообразных может прожить от 3 до 4 дней, а питается он в это время желтком, который находится в желточном мешочке на спине под щитком.
Так маленький представитель ракообразных может прожить от 3 до 4 дней, а питается он в это время желтком, который находится в желточном мешочке на спине под щитком.
https://youtube.com/watch?v=6bzLZQsYeas
Первая линька малышей
Четверо суток после появления на свет личинка имеет мягкий панцирь. На шестой день наступает первая в её жизни линька: желточный мешок пропадает, панцирь твердеет, личинка съедает яйцевую оболочку. Через неделю малыш становится очень активным и начинает плавать возле самки. В возрасте двух недель может навсегда покинуть её и отправиться в самостоятельную жизнь.
Период линьки очень опасен для раков. Сбрасывается не только панцирь, но и покрытие жабр, глаз, пищевода и зубов, которыми они поедают пищу. Твёрдыми остаются только гастролиты, которые располагаются в желудке и накапливают кальций, необходимый для образования твёрдых частей тела. Рак, который линяет, становится беззащитным, и поэтому почти весь период старается не выползать из своей норы, чтобы не стать лёгкой добычей хищников или сородичей-каннибалов.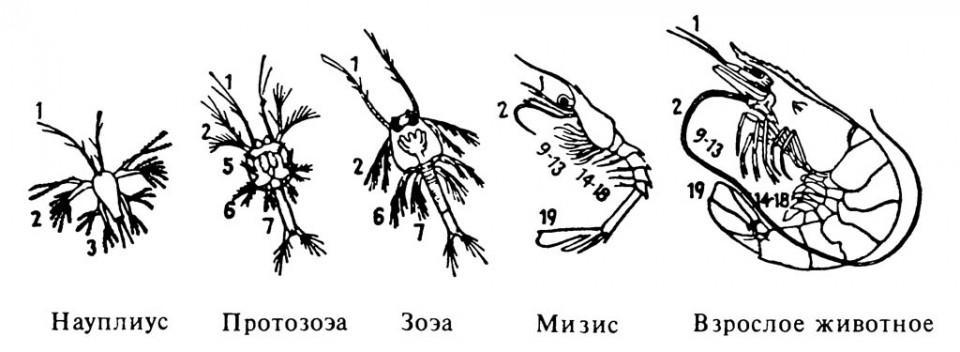
Бизнес на разведении
Членистоногих разводят не только в естественных водоёмах. Аквариум — один из способов размножения речных раков.
В домашних условиях преимущественно разводят такие виды:
- Голубой кубинский. Вид популярен своим быстрым ростом и всеядностью. Эти раки спариваются при температуре не выше +26°С.
- Австралийский рак считается мясным видом, который хорошо разводится в аквариумах.
- Мраморный рак относится к гермафродитам. При его выращивании нужны большие пространства и постоянная температура воды.
Особей для размножения можно приобрести в универсамах или наловить самому. Лучшим вариантом станет приобретение личинок. Объем аквариума потребуется не меньше 250 литров. Имитируя природную среду, на дно укладывают камни, коряги, песок.
В аквариум для спаривания и получения молодой поросли запускают 3 взрослых особей.
Их плотность в ёмкости, чтобы они могли размножаться, не должна превышать 350 штук на один квадратный метр.
Уход и подкормка
Аквариумное размножение раков требует много времени. Выращиваемые в аквариумах, они не нуждаются в спячке и намного быстрее набирают вес.
При разведении членистоногих в аквариумах следует соблюдать такие условия:
- постоянная температура;
- чистая вода, снабжённая кислородом;
- сбалансированный корм;
- обязательная витаминная подкормка.

Рак — травоядное существо, но в природных условиях в его рацион входит падаль. При недостаче еды они пожирают своих сородичей.
В неволе членистоногих кормят варёными зерном и картофелем, сырым мясом, тёртой морковью, рыбой, личинками червей, насекомых, улиток, специальными кормами. В природных условиях в их рацион попадают зелень, дождевые черви, улитки, мелкая рыба. Клешнями членистоногие захватывают добычу, отрывают от неё кусочки и съедают их.
Разведение раков | Справочник аквариумиста
Аквариумные раки при создании благоприятных условий размножаются так же активно, как и аквариумные креветки. Размножение раков будет происходить без негативных последствий, если содержание и кормление раков в аквариуме будет отвечать их естественным потребностям. Получение потомства необходимо для поддержания поголовья карликовых раков в аквариуме, так как продолжительность жизни этой группы раков варьируется от 14 до 30 месяцев в зависимости от вида.
Размножение раков будет происходить без негативных последствий, если содержание и кормление раков в аквариуме будет отвечать их естественным потребностям. Получение потомства необходимо для поддержания поголовья карликовых раков в аквариуме, так как продолжительность жизни этой группы раков варьируется от 14 до 30 месяцев в зависимости от вида.
Определение пола у раков. Определить пол у раков не так просто. В зависимости от того, самец это или самка, половые отверстия расположены в разных частях тела. Половые отверстия самцов, которые называют трубчатыми, расположены у основания последней пары ходильных ног. Самки имеют половые отверстия (гоноподы) перед третьей (если считать от последней) парой ног. Между прочим, многие особи имеют признаки как мужских, так и женских половых отверстий, делая определение пола затруднительным. Тем не менее, раки всегда однополы: являются или самцами, или самками.
Внешне взрослых мужских и женских особей можно различить по клешням и хвосту. У самцов клешни более длинные и крупные, у самок же шире хвост. Так же как и аквариумные креветки, аквариумные раки носят икру под брюшком, поэтому эта часть тела у самок шире. Головогрудь и брюшко самок шире, чем самцов, клешни тупые, короткие. У самок первые брюшные ножки либо отсутствуют совсем, либо значительно меньше по размеру. Плодовитость большинства декоративных декапод существенно зависит от размеров самки. Чем она крупнее и соответственно старше, тем больше яиц в состоянии носить.
У самцов клешни более длинные и крупные, у самок же шире хвост. Так же как и аквариумные креветки, аквариумные раки носят икру под брюшком, поэтому эта часть тела у самок шире. Головогрудь и брюшко самок шире, чем самцов, клешни тупые, короткие. У самок первые брюшные ножки либо отсутствуют совсем, либо значительно меньше по размеру. Плодовитость большинства декоративных декапод существенно зависит от размеров самки. Чем она крупнее и соответственно старше, тем больше яиц в состоянии носить.
В период размножения раки становятся очень активными. В поисках самки самцы уходят далеко от убежищ и даже днем ползают по дну водоема. Если в это время поставить аквариум с самкой вблизи от аквариума с самцом, тот проявляет явное беспокойство, встает на дыбы и стучит клешнями о стекло, отделяющее его от самки. Самки многих видов раков готовы к спариванию и откладыванию яиц лишь в течение короткого периода после линьки (правда, есть и исключения). В это время они активно выделяют особые вещества — феромоны, предназначенные для привлечения полового партнера.
Чаще всего эти аттрактанты видоспецифичны, то есть, адресованы лишь самцам-сородичам, но порой могут заинтересовать и мужскую половину близкородственных видов. Период гона высших раков обычно заставляет нервничать начинающих любителей беспозвоночных. Им невдомек, почему прежде спокойные питомцы внезапно начинают бешено носиться по всему аквариуму. На самом деле ничего страшного не происходит, просто раздраженные призывными запахами самцы активно ищут партнершу.
Стимуляция размножения. Для того чтобы раки начали размножаться, необходимо создать для этого подходящие условия. Места в аквариуме должно быть достаточно (на одного взрослого рака 15-20 литров или ориентировочно 30 х 40 см площади дна), питание должно быть разнообразным и полноценным. У некоторых видов раков стимуляторами к размножению являются изменения внешних условий окружающей среды — изменение количества световых часов, температуры воды или циркуляции воды.
В природе ливни и вызванный ими приток свежей воды являются для десятиногих ракообразных сигналом к началу размножения, а удлинение светового дня и повышение температуры стимулируют созревание половых продуктов. Регулярная смена части воды на свежую, стимулирует половозрелость раков. Слейте часть воды из емкости шлангом и долейте взамен водопроводную той же температуры или на несколько градусов холоднее.
Регулярная смена части воды на свежую, стимулирует половозрелость раков. Слейте часть воды из емкости шлангом и долейте взамен водопроводную той же температуры или на несколько градусов холоднее.
Раки питаются пищей как растительного, так и животного происхождения. Чтобы поддерживать их в добром здравии и отличной форме, рекомендуется давать им вареный салат, мясо рыбы или креветок (раз в неделю) или специализированный корм для раков. Такое питание поможет самке окрепнуть перед вынашиванием икры и продержаться в период беременности без еды, так как обычно самки в этот период ничего не едят.
Спаривание и вынашивание икры. Аквариумные раки в отличие от диких размножаются круглый год. Самец, уложив самку на спину и держа ее за клешни своими клешнями, помещает сперматофоры (капсулы со спермой) на живот самки в районе половых отверстий. Самка, получившая от самца пакет спермы, перестает вырабатывать феромоны и больше не представляет интереса для особей противоположного пола. Спермопакет прикрепляется непосредственно у полового отверстия самки. Оплодотворение наступает в тот момент, когда яйца покидают ее тело и по пути к плеоподам проходят через спермопакет.
Спермопакет прикрепляется непосредственно у полового отверстия самки. Оплодотворение наступает в тот момент, когда яйца покидают ее тело и по пути к плеоподам проходят через спермопакет.
После оплодотворения яйцеклеток спермой, самка откладывает икру на плеоподы (брюшные ноги). Для их фиксации на брюшных ножках и крепления яиц между собой служат так называемые нити-тяжи. Самка с икрой передвигаться осторожно, часто прячется в укрытиях. При опасности поджимает хвост, пряча икру. Теперь ее главная забота — вентиляция кладки, удаление неоплодотворенных или мертвых яиц и эмбрионов. Постоянно находящиеся в движении плавательные ноги самки обеспечивают вентиляцию кладки, доставку к яйцам кислорода.
Длительность периода вынашивания икры зависит от температуры воды и вида раков и занимает не менее 3 недель. На продолжительность вынашивания икры могут влиять и другие факторы, такие как качество воды и корма. В этот период крайне необходимо обеспечить самку надёжным укрытием, которое обеспечит безопасность самки и ее потомства. Прекрасно для этого подходят укрытия, сделанные из скорлупы кокосового ореха. Идеальным вариантом была бы отсадка беременной самки в отдельный аквариум, где ее не потревожат ни самцы, ни крупные рыбы. Во время вынашивания икры не рекомендуется тревожить самку и производить большую подмену воды.
Прекрасно для этого подходят укрытия, сделанные из скорлупы кокосового ореха. Идеальным вариантом была бы отсадка беременной самки в отдельный аквариум, где ее не потревожат ни самцы, ни крупные рыбы. Во время вынашивания икры не рекомендуется тревожить самку и производить большую подмену воды.
Рачата. Новорожденные рачки очень мелкие, но, тем не менее, они представляют собой полностью сформированных раков. Размер новорожденных раков около 7-9 мм. Вылупляющиеся рачки сначала неподвижно висят на ножках самки, а после первой линьки пробуют от нее уходить, но в минуту опасности быстро возвращаются и прячутся ей под брюшко. Стартовым кормом для малька служит то же, что едят взрослые. А также различные органические остатки, которых обычно много в аквариуме. Выкармливать молодь можно готовыми кормами для мальков рыб, резаным трубочником, артемией.
Как правило, маленькие рачки – детритофаги, то есть питаются разлагающимися частичками растений и остатков корма, которые они находят на дне аквариума. В первые недели молодь ищет себе пищу недалеко от матери. Как только самка начинает перемещение по аквариуму или рачата чувствуют опасность, тут же забираются матери под брюшко. Лишь после второй линьки они начинают активно двигаться и питаться, покидают мать и переходят к самостоятельному образу жизни.
В первые недели молодь ищет себе пищу недалеко от матери. Как только самка начинает перемещение по аквариуму или рачата чувствуют опасность, тут же забираются матери под брюшко. Лишь после второй линьки они начинают активно двигаться и питаться, покидают мать и переходят к самостоятельному образу жизни.
По своей природе раки агрессивны друг к другу, борьба за территорию часто приводит к смерти противника. И этот инстинкт проявляется с раннего возраста. Обитание рачков в маленьком аквариуме может привести к каннибализму. По мере подрастания выводка необходимо предоставлять ракам более просторные емкости, т.к. в противном случае в маленьком объеме они будут наносить друг другу повреждения, вплоть до поедания друг друга. В природе от дефектов линьки погибает довольно много рачьей молоди. Так что даже при идеальном содержании потерь не избежать. Сбросив шкурку, они становятся легкой добычей не только хищников, но и своих сородичей.
Поэтому если есть желание поднять как можно большее количество молоди раков, необходимо позаботиться о просторном аквариуме, в котором основную роль играет площадь дна и наличие укрытий в виде камней и керамических черепков. Поскольку в первые недели молодняк часто линяет, следует позаботиться о наличии достаточного количества корма и укрытий. В противном случае не избежать больших потерь из-за каннибализма. При разведении раков хорошо зарекомендовали себя полые строительные кирпичи. Они не только дешевы, но и обеспечивают юных декапод большим количеством убежищ. Молодняку нужно предоставить в распоряжение густо заросший аквариум.
Поскольку в первые недели молодняк часто линяет, следует позаботиться о наличии достаточного количества корма и укрытий. В противном случае не избежать больших потерь из-за каннибализма. При разведении раков хорошо зарекомендовали себя полые строительные кирпичи. Они не только дешевы, но и обеспечивают юных декапод большим количеством убежищ. Молодняку нужно предоставить в распоряжение густо заросший аквариум.
(Visited 2 786 times, 5 visits today)
Выращивание речных раков — Агропромышленный портал Агро-Спутник
Лидером по выращиванию речных раков является Турция, кудав свое время они были завезены из водоемов Украины. Ежегодно, на внешний рынок, Турция поставляет до 7 тыс. т товарных раков, несколько меньше Испания – 3,5 тыс. и Китай – 1 тыс. т.
Активно выращивать раков начали в Иране. Примечательно, что местные жители таких стран, как Турция и Иран, не используют раков в пищу, вся выращенная продукция идет на экспорт. В европейских странах, например Германии и Франции, выращивают широкопалого и длиннопалого рака. В России наиболее распространен длиннопалый рак.
В России наиболее распространен длиннопалый рак.
Раки всеядны. В большинстве случаев они не являются конкурентами рыбе. Поедая останки погибших животных, раки являются «санитарами водоемов».
Самцы длиннопалых раков становятся половозрелыми на третий год жизни при длине тела 8 см, а самки – только на четвертый год при длине тела 7 см, т.е. самцы, как правило, крупнее самок. Спаривание у раков происходит либо осенью (октябрь-ноябрь), либо в конце зимы – начале весны (февраль-март). Спаривание продолжается в течение двух-трех недель, оплодотворение при этом внешнее. Самцы приклеивают свои сперматофоры на нижней стороне головогруди самки в форме белого пятна. При позднем спаривании и наступлении холодов, оплодотворение длится несколько дней.
Самки откладывают икру ночью, на протяжении 2–3 ч. Они подгибают брюшко к головогруди, образуя камеру, куда выпускают специальное вещество, растворяющее прикрепленные там сперматофоры со сперматозоидами. Затем икринки выходят из яйцевода, проходят через семенной раствор, оплодотворяются и прикрепляются к брюшным ножкам или панцирю. Самка может вынашивать до 100–150 икринок, ее плодовитость зависит от возраста, массы и состоя- ния. Период развития эмбрионов в икре составляет 50–60 суток.
Самка может вынашивать до 100–150 икринок, ее плодовитость зависит от возраста, массы и состоя- ния. Период развития эмбрионов в икре составляет 50–60 суток.
Вылупление личинок раков происходит весной или летом при температуре 21–24°С. В возрасте десяти дней происходит первая линька, после которой личинка становится похожей на взрослых раков. При этом масса ее тела составляет 21–30 мг, длина – 1,1–1,2 см. Рачки начинают питаться, но при необходимости прячутся под брюшко матери. И только на 15–20 день после второй линьки, молодые раки становятся полностью самостоятельными. Личинок, на первой стадии развития, кормят мелким зоопланктоном, на последующих стадиях – искусственным кормом, например, стартовым карповым или форелевым комбикормом, измельченной рыбой, мясом, отходами боен. Полноценное питание снижает отход молоди, так как у личинок проявляется каннибализм уже после второй линьки. После пяти линек, т. е. к концу августа, молодь достигает длины 2–2,5 см, к этому времени спектр их питания расширяется. Они начинают потреблять растительную пищу. Характерными кормовыми участками водоема являются прибрежное мелководье (вплоть до уреза воды), места, заросшие водной растительностью. Успех выращивания раков во многом химического состава воды и характеристики грунтов водоема.
Они начинают потреблять растительную пищу. Характерными кормовыми участками водоема являются прибрежное мелководье (вплоть до уреза воды), места, заросшие водной растительностью. Успех выращивания раков во многом химического состава воды и характеристики грунтов водоема.
Раки предпочитают обитать на галечно-песчаных, глинисто-песчаных грунтах с небольшой примесью не гниющего органического ила. Наиболее интенсивно раки растут при температуре воды – 18–22 °С. Они предпочитают воду низкой (3–4 мг экв/л) и средней (4–8 мг экв/л) жесткости при слабощелочных значениях рН (7,2–8,8). Критическим значением рН воды для раков является показатель 4,6. Содержание ионов сульфата и хлорида не должно превышать показатель 10 мг/л, а общего железа – 0,3 мг/л.
Раки особенно уязвимы в молодом возрасте, так как ими питаются рыбы (щука, сом, сазан), водяные млекопитающие, водоплавающие птицы, ужи. В этом возрасте, раки предпочитают питаться беспозвоночными (личинки насекомых, черви, мелкие ракообразные, водные насекомые, рыбы), потребляют в пищу умершую рыбу, но, преимущественно, свежую. Кроме того, ракам необходимо потреблять водоросли, являющиеся источником кальция, который формирует изоскелет.
Кроме того, ракам необходимо потреблять водоросли, являющиеся источником кальция, который формирует изоскелет.
Пол взрослых раков определяют по размеру клешней и хвосту: у самцов клешни бoльшего размера, а у самок шире хвост, который защищает икру во время всего периода ее развития под хвостом. Пол молодых раков визуально можно установить только по наличию или отсутствию у них трубчатых половых органов. Половое отверстие у самок находится у основания третьей пары конечностей, а у самцов – четвертой пары.
Линька раков связана с быстрым ростом тела: тело перестает умещаться в старую оболочку, требуется ее замена. Раки становятся половозрелыми в возрасте 3–4 лет, можно ожидать появления полноценного самовоспроизводящегося стада примерно через 4–5 лет. С этого момента можно начинать их отлов, но не более 20% взрослых раков в год, чтобы не нарушить сложившуюся структуру стада.
Крупными раки (длиной 18–20 см, массой 250–300 г) становятся на шестом-девятом году жизни.
Существует множество авторских технологий выращивания раков. Основное условие для выращивания раков – наличие водоемов с малозаиленным глинистым дном и водой, обогащенной кислородом, поддержание постоянного летнего температурного и гидрохимического режимов.
Основное условие для выращивания раков – наличие водоемов с малозаиленным глинистым дном и водой, обогащенной кислородом, поддержание постоянного летнего температурного и гидрохимического режимов.
Для разведения раков можно использовать специальные пруды, фермерские водоемы, а также небольшие частные и приусадебные участки, на которых имеются водоемы. Различают две разные технологии разведения раков – прудовую и заводскую (индустриальная). Первая является более выгодной, так как раки требуют собственное, индивидуальное место – укрытие, которое легче найти в пруду. Им противопоказана кислая водная среда и загрязнение водоемов любыми сточными водами. Раки предпочитают воды с достаточным содержанием кислорода (6–8 мг/л). При прудовой технологии выращивания и разведения раков отловленных производителей высаживают в специально подготовленные спускные пруды площадью 0,1 га, имеющие мелководную и глубокую части. В прудах, куда помещают раков в конце лета, происходит естественный нерест. Плотность посадки 1–5 шт/м2. При повышении температуры воды более 7°С их начинают кормить, задают свежий или вареный корм (отходы боен, овощи, рыба, моллюски и т.п.), при этом средняя суточная норма выдачи корма должна составлять 2% от массы тела рака. Влажные корма размещают на деревянных лотках (40×40 см), раки также питаются сухими кормовыми гранулами.
Плотность посадки 1–5 шт/м2. При повышении температуры воды более 7°С их начинают кормить, задают свежий или вареный корм (отходы боен, овощи, рыба, моллюски и т.п.), при этом средняя суточная норма выдачи корма должна составлять 2% от массы тела рака. Влажные корма размещают на деревянных лотках (40×40 см), раки также питаются сухими кормовыми гранулами.
При прудовом способе разведения раков личинки вылупляются в мае-июне. За одно лето раки достигают возраста сеголеток массой 7–10 г, которых оставляют в этом пруду на зимовку, если глубина пруда более 1,5 м, или пересаживают в другие пруды. Следующей весной годовиков отлавливают и пересаживают в нагульные пруды с меньшей плотностью посадки. В конце второго или на третьем году жизни раки достигают товарной массы (40–60 г) при длине 9–10 см. Зимой раки уходят на глубину и большую часть времени проводят в норах. Даже в условиях пониженной температуры, раки не прекращают активно питаться. Чтобы получить 3–4 ц/га товарной продукции речных раков, необходимо иметь не меньше трех-четырех прудов.
Материал подготовлен специалистами ГБУ КК «Кубанский сельскохозяйственный ИКЦ»
Газета «Аграрная Кубань»
15 августа 2012 г.
рацион в природе и домашних условиях
Речные раки довольно часто становятся питомцами, содержащимися в домашних условиях в искусственных водоемах. К сожалению, большинство аквариумистов до сих пор полагает, что в качестве пищи им достаточно падали, водорослей, остатков корма для рыб и продуктов их жизнедеятельности. На самом же деле рацион речных раков куда разнообразнее и от него зависит здоровье, а также продолжительность жизни питомцев.
Особенности питания речных рачков
Половая принадлежность. Количество пищи и рацион речных рачков зависят от половой принадлежности. Самцы питаются ежедневно, принимая пищу 1 раз на протяжении суток. А самочки едят лишь 1 раз в 3 дня, но употребляют значительно большие по объему порции.
Время для кормления. Речные рачки ведут ночной образ жизни. Поэтому аквариумистам, содержащим таких питомцев в домашних условиях, следует кормить животных в вечернее время суток.
Поэтому аквариумистам, содержащим таких питомцев в домашних условиях, следует кормить животных в вечернее время суток.
Объем порции. Определить необходимое количество корма − простая задача. Речные рачки не склонны к перееданию. Поэтому достаточно просто поставить в аквариум кормушку, наполнив ее кормом. Как только емкость окажется пустой, в нее следует положить новую порцию пищи.
Однако накладывать слишком большие порции не рекомендуется. Остатки пищи способствуют загрязнению аквариума, поэтому его придется очищать как минимум 1 раз на протяжении 2 суток, чтобы избежать развития гнилостных процессов, отрицательно сказывающихся на здоровье аквариумных обитателей.
Чем питаются речные рачки в природных условиях
Ракообразные отдают предпочтение растительной пище, лишь 10% их рациона составляют животные корма.
В природной среде обитания речные рачки используют в качестве корма:
- фитопланктон;
- водоросли;
- органику;
- мелких беспозвоночных существ.

Сезонность. Но их вкусовые предпочтения зависят от времени года и ряда других факторов. Например, в период размножения или зимнюю пору речные раки отдают предпочтение белковой пище в виде червей, головастиков, личинок, моллюсков, мелких рыбешек.
Вопреки ошибочно распространенному мнению, речные рачки не питаются падалью. Вернее, они могут употреблять ее в пищу, но лишь в случае крайней необходимости, при отсутствии какой-либо другой пищи.
В теплое время года рацион питания речных раков, которые могут существовать как в водной среде, так и на суше, значительно расширяется.
Ракообразные с огромным удовольствием употребляют в пищу стебли, листовые пластины, корни различных растений.
Особенности кормления речных рачков в домашних условиях
Рацион речных рачков в природе и при содержании в искусственной среде заметно отличаются. Аквариумистам необходимо знать, чем и как кормить своих питомцев, чтобы уберечь их от ряда заболеваний и быстрой гибели.
Речные рачки − всеядны, поэтому проблем с выбором корма, как правило, не возникает. В рацион питомцев можно включать:
- хлебные крошки;
- измельченные кусочки мяса любых сортов;
- злаки;
- различные каши;
- кусочки свежих овощей и фруктов;
- зелень;
- крапиву;
- дождевых червей.
Кроме того, можно приобрести специализированные корма, предназначенные именно для ракообразных, которые продаются в любом зоомагазине.
Отлично подойдут для речных рачков такие виды замороженного или живого корма, которые используются для кормления декоративных рыбок, например:
- мелкие моллюски;
- циклопы;
- дафния;
- коловратка.
Белковая пища также необходима ракообразным, но основу их рациона все же должны составлять растительные корма.
Самое главное − это следить за порциями предлагаемого корма.
Специалисты рекомендуют давать корм в объеме, не превышающем 3% от массы тела речного рачка.

В противном случае остатки не съеденной пищи начинают гнить, разлагаться, что приводит к гибели членистоногих.
Природные корма для речных рачков
Облегчить задачу, связанную с кормлением речных раков, вполне можно, заранее обустроив искусственный водоем, предназначенный для проживания данных питомцев. Для этого достаточно высадить на днище аквариума:
- люпин;
- бобовые;
- горох;
- овес.
Эти растения благотворно сказываются на состоянии водной среды, насыщая ее и почву оптимальным количеством азота. А самое главное − речные раки могут использовать их в качестве корма.
Читайте также:
Зависимость рациона питания от возраста речных рачков
Важно учитывать, что вкусовые предпочтения и потребность в определенной пище у речных рачков меняется по мере их роста и развития. Специалисты разработали определенную схему кормления ракообразных на разных этапах их жизни:
- Сеголетки − питаются преимущественно дафнией.
- Раки, достигшие размеров до 2 см − нуждаются в белковой пище. Подкармливать их можно личинками. В этот период животный корм должен составлять около 45% от общего рациона.
- Рачки размером от 3 см − могут употреблять в пище моллюсков и насекомых.
- Молодняк размером от 4 см − включают в свое меню мелкую рыбу.
- Взрослые особи (8-10 см) − питаются преимущественное бокоплавом, отдавая предпочтение растительной пище.
Перечисленные выше особенности следует учитывать при выращивании раков в искусственном водоеме, что особенно важно для аквариумистов, занимающихся разведением и размножением данных животных. Дополнив рацион речных рачков пищей, которую они употребляют в природной среде обитания, можно быть уверенным в их здоровье, активном росте, полноценном развитии.
Речные рачки в последнее время часто становятся обитателями искусственных водоемов. Несмотря на неприхотливость данных существ в плане еды, важно следить за их рационом и соблюдать определенные правила кормления, которые отличаются от питания моллюсков или декоративных рыбок. Необходимо обеспечить речным рачкам грамотное питание с преобладанием растительной пищи в рационе и дополнительными белковыми добавками в виде живых или замороженных кормов, мяса, морепродуктов.
Необходимо обеспечить речным рачкам грамотное питание с преобладанием растительной пищи в рационе и дополнительными белковыми добавками в виде живых или замороженных кормов, мяса, морепродуктов.
Рубрики: Чем кормить |
автор Olga
уход, содержание, размножение, совместимость, корм, фото-обзор
Аквариумные обитатели с желтой или оранжевой окраской тела необычайно популярны. И дело здесь не только в том, что окрас сразу бросается в глаза. Подобные цвета придают аквариуму нотки тепла, света, энергии, многократно усиливая релаксирующий эффект. В нашей статье мы расскажем вам об одном аквариумном «солнышке» – мексиканском карликовом раке.
Общие сведения
Мексиканский
карликовый рак (Cambarellus patzcuarensis var. Orange) – пресноводное ракообразное из
отряда Десятиногие раки. Также можно встретить обозначение по первым буквам
латинского названия – рак CPO.
Этот житель Центральной Америки отличается миниатюрными размерами,
неприхотливостью в уходе и кормлении, а также потрясающей оранжевой или красной
окраской тела. Все это делает его идеальным кандидатом для содержания в
домашних аквариумах.
Все это делает его идеальным кандидатом для содержания в
домашних аквариумах.
Популярности добавляет и тот факт, что рака не обязательно содержать в видовом аквариуме: он неплохо ладит как с некоторыми популярными рыбками, так и с живыми растениями.
Мексиканский карликовый рак красив и миролюбивСтоит отметить, что прекрасная оранжевая «шкурка» мексиканского карликового рака – это результат работы селекционеров, ведь в природе подобные цвета сделают из него легкую добычу для хищников. В аквариуме же ему бояться некого, поэтому раки продолжают радовать нас яркими нарядами.
Характерны
для раков и интересные формы поведения: «обустройство» своего убежища,
регулярные линьки, спаривание и вынашивание потомства, стычки за территорию. А
чего стоит момент, когда рак голоден – он бегает вдоль стенки аквариума, подняв
свои клешни. Эти питомцы любят повисеть вниз головой, зацепившись за стебли
растений, такой «ритуал» может длиться несколько часов и порой приводит в
недоумение начинающих раководов.
Мексиканские карликовые раки ведут дневной образ жизни, что позволяет легко наблюдать за их существованием в домашнем аквариуме. Редко какое аквариумное животное приносит такую бурю эмоций и удивления.
Внешний вид
Если вы хоть раз видели обыкновенного рака, то представить себе внешний вид героя статьи вам будет несложно. Тело состоит из характерных для ракообразных отделов: плотной головогруди и подвижного брюшка, оканчивающегося широким хвостовым плавником. Первый отдел служит местом закрепления ходильных ног (пять пар, первая заканчивается массивными щупальцами), челюстей и органов чувств. Под плотным панцирем (карапаксом) под надежной защитой находятся внутренние органы рака. Брюшко состоит из сочлененных сегментов, под которыми расположены плавательные ножки – плеоподы. Они участвуют не только в плавании, но и в размножении. Например, у самца одна из пар превратилась в орган копуляции, а самки используют их для вынашивания икры и молодых рачат.
Мексиканский карликовый рак. Внешний видНаиболее популярная окраска тела – красно-оранжевая, интенсивность которой может варьировать. Вдоль головы и хвоста могут располагаться темные или светлые полосы, а также небольшие округлые пятна. Длина тела не превышает 5 см.
Продолжительность жизни в аквариумных условиях составляет 2-3 года.
Ареал обитания
Как несложно догадаться из названия, родиной карликового рака является территория Мексики. Одно из наиболее известных мест распространения – вулканическое озеро Лаго-де-Пацкуаро (мексиканское побережье Тихого океана), которое распложена на высоте 2000 м над уровнем моря, поэтому вода в нем остается относительно прохладной даже летом.
Живут раки преимущественно в стоячих водоемах, либо же в ручьях и речках с несильным течением. Держатся возле берегов, ведь именно здесь проще всего найти подходящие укрытия среди корней деревьев или горных пород береговой линии.
Уход и содержание
Для содержания пары мексиканских карликовых раков понадобится аквариум от 50 литров. Лучше всего, если у него будет максимальная площадь дна, ведь именно здесь проходит почти вся жизнь раков.
В качестве грунта подойдут крупный песок или средняя галька. Слой лучше сделать потолще, поскольку раки очень любят перекапывать дно.
В аквариуме с мексиканскими карликовыми раками должно быть большое число разнообразных укрытий. Это поможет вашим питомцем разделить территорию, если в емкости живет несколько особей, прятаться во время линьки, когда незатвердевший покров очень уязвим, а самкам еще и сохранять потомство. В качестве укрытий подойдут любые подручные материалы: гроты, керамические горшки, полые трубки диаметром 2-5 см, скорлупа кокоса и т.п. Нередко раки устраивают возле входа в убежище импровизированную дверь – нагребают горку грунта, которая закрывает вход, когда покидают дом, раскапывают ее, а по возвращении снова зарывают.
Мексиканский карликовый рак в аквариумеАквариум должен быть обязательно оборудован фильтром. Это необходимо для поддержания чистоты и высокого качества воды, ведь раки чувствительны к высоким содержаниям азотистых соединений. Предпочтение лучше отдать внутренним фильтрам. Это связана с тем, что мексиканские раки – отличные «альпинисты». Они с легкостью могут взобраться по трубкам внешнего фильтра и совершить побег через технологические отверстия. К сожалению, такое путешествие будет фатальным для рака, если его вовремя не вернуть в аквариум, так как без воды они долго прожить не смогут. Особое внимание необходимо уделить аэрации. Для раков жизненно важен растворенный в воде кислород, поэтому стоит выбирать фильтры с подачей воздуха, а еще лучше установить отдельный компрессор. Особенно актуальна проблема нехватки кислорода в летний период, ведь в теплой воде он растворяется гораздо хуже.
Мексиканские карликовые раки хорошо относятся к живым растениям. Единственная угроза – вероятность подкопа корней. Чаще всего рекомендуются неприхотливые виды растений: пучки яванского мха, роголистник, криптокорина, папоротники. На поверхность воды желательно запустить пистию или риччию – они создают естественный полумрак, так как раки не любят яркого освещения.
В аквариумах с мексиканскими карликовыми раками должно быть много укрытийРаз в неделю следует подменивать 20% воды в аквариуме на свежую, при этом водопроводную воду обязательно следует подготовить с помощью кондиционера Tetra Crusta AquaSafe, который удалит из воды опасные для раков соединения.
Вода для содержания раков не должна быть слишком мягкой, так как членистоногим требуются минералы для формирования панциря.
Оптимальные параметры воды для содержания: Т=18-25°С, pH=7.0-8.5, GH=10-20.
Одной из особенностей в содержании раков является их периодическая линька. Она заключается в сбрасывании старого покрова, который уже не позволяет раку расти, и формировании нового, в этот период ракообразные увеличиваются в размерах, пока панцирь не затвердеет. В течение этого времени мексиканские карликовые раки являются наиболее уязвимыми, ведь без защитного покрова их способны обидеть даже миролюбивые виды рыб, не говоря уже о своих собственных сородичах. Линька позволяет также отрастить новые конечности, утраченные в боях при групповом содержании раков. Очень важно не выбрасывать сброшенный покров из аквариума, он будет съеден хозяином, а его питательные элементы пойдут на построение новой «одежки».
Совместимость
Мексиканские карликовые раки отличаются миролюбивым характером. Внутривидовая агрессия имеет место быть, но она стремится к нулю при отсутствии перенаселения и большом количестве разнообразных укрытий. В таких ситуациях даже возможные драки оканчиваются благополучно.
С рыбами у раков отношения спокойные. Чаще всего бывает, что подвижные соседи являются дополнительным источником стресса для раков, которые при их появлении стараются как можно быстрее спрятаться в убежище. Даже длительное сожительство с одними и теми же видами не делает членистоногих менее боязливыми.
С другой стороны, голодный рак будет, словно танк, продвигаться к заветному корму, расталкивая даже рыб крупнее себя.
Мексиканский карликовый рак хорошо уживается с рыбками и живыми растениямиЛучше всего для совместного содержания подойдут подвижные рыбки среднего размера: моллинезии, пецилии, барбусы. С донными рыбками отношения также обычно складываются хорошие, несмотря не необходимость делить территорию, поэтому коридорасов, торакатумов, анциструсов, боций можно селить с раками без боязни.
Из нежелательных видов можно выделить различных цихлид (чернополосая цихлазома, северум, акара, астронотус и др.), а также креветок и крабов.
Кормление мексиканского карликового рака
Питание мексиканского карликового рака – самая малая из проблем при содержании. Эти животные совершенно всеядны, происхождение пищи для них не играет никакой роли. Отмершие части растений, погибшая рыба, сухие корма – все это будет съедено раком с удовольствием. Широко распространенные живые и замороженные корма лучше не использовать, они не содержат всего комплекса питательных веществ, необходимых раку, а вдобавок могут очень быстро испортить воду в случае, когда не съедаются питомцами полностью.
Лучше всего остановиться на качественных сухих кормах для ракообразных Tetra Crusta. Эти полностью сбалансированные корма обеспечат рака всем необходимым, в том числе питательными элементами для хорошей линьки. Они быстро опускаются на дно и длительное время сохраняют свою форму, что позволяет кормить раков даже в общих аквариумах. При этом корм не вызывает помутнения воды и ухудшения ее качества. Также раки отлично будут поедать универсальные корма для сомов и донных рыб – Tetra WaferMix или Tetra Pleco SpirulinaWafers.
Дополнительно можно подкармливать рака высушенными листьями дуба или индийского миндаля.
Размножение и разведение
Получить потомство от мексиканского карликового рака в домашних условиях проще простого. Вам понадобится пара производителей и небольшой нерестовый аквариум литров на 20-30.
Половая зрелость у раков наступает очень рано – в 3-4 месяца. Отличить самца и самку достаточно легко. Прежде всего, обратите внимание на клешни: у самки они короткие и толстые, хотя сама она крупнее самца. У мужских же особей клешни вытянутые. Для подтверждения информации возьмите рака и поверните его на спину, после чего взгляните на первую пару плеоподов – у самца она ланцетовидная, длинная (орган копуляции), а у самки короткая.
Мексиканские карликовые раки легко разводятся в домашних условияхПосле оплодотворения икра развивается у самок от одной до четырех недель. Затем она прячется в укрытие, где появляются икринки и тут же прочно прикрепляются к плавательным ножкам матери. Инкубация икры займет срок до месяца-полтора. Все это время самка практически не выходит из укрытия, а с помощью плеоподов создает постоянный ток воды возле икринок. Плодовитость женской особи может составлять до 60 икринок. В период вынашивания икры корм самке стоит бросать прямо перед входом.
Оплодотворенную самку лучше всего отсадить в нерестовый аквариум, который должен быть с небольшим аэрлифтным фильтром, а на дно положить пучок мелколистных растений (роголистник, яванский мох и т.п.), а также создать множество укрытий. Они понадобятся молодым рачкам, чтобы потери потомства от каннибализма были минимальны.
Появившиеся на свет рачки являются точными копиями своих родителей, только в уменьшенном размере, первое время они держаться за плеоподы матери, но вскоре начинают самостоятельную жизнь.
Первое время молодые раки очень часто линяют (из-за усиленного роста), но с возрастом этот процесс происходит все реже и реже.
Научное название: Cambarus sp . (Информация на этой странице видов была собрана Алисией Фицджеральд в журнале Biology 220W, Spring 2006, в Пенсильвании, Нью-Кенсингтон) Раки (с особым упором на род Cambarus ) — чрезвычайно характерные пресноводные ракообразные.Их большие передние пары ног имеют мощные когти, которые являются эффективными инструментами для защиты, сбора пищи и манипулирования объектами. Их четыре пары ходильных ног помогают им быстро перемещаться по дну их водной среды обитания. Удивительно, но они на самом деле не умеют плавать, хотя могут использовать мощные толчки хвоста, чтобы быстро продвигаться (назад) по воде. Последнее движение особенно эффективно, когда рак пытается ускользнуть от хищника. Распределение Пределы экологических допусков Диета Линия Спаривание и размножение Хищники, паразиты, симбиоз и болезни Экологическое значение |
(PDF) Рост и размножение раков
114 Биология и экология раков
Momot, W.T. 1984. Производство раков: отражение энергетики сообщества. J. Crust. Биол. 4: 35–54.
Morgan, G.J. 1997 г. Пресноводные раки из рода Euastacus Clark (Decapoda: Parastacidae) из Нового Южного Уэльса, при этом
являются ключом ко всем видам этого рода. Рек. Aust. Mus. 23: 1–110.
Muck, J.A., C.F. Рабени и Р.Дж. Distefano. 2002. Репродуктивная биология раков Orconectes luteus (Creaser) в ручье
Миссури. Являюсь. Midl. Nat. 147: 338–351.
Наката, К.и С. Госима. 2004. Плодовитость японского рака Cambaroides japonicus: формирование завязи, число яиц
и размер яиц. Аквакультура 242: 335–343.
Наката, К., А. Танака и С. Госима. 2004. Размножение чужеродного ракового вида Pacifasticus leniusculus в озере
Сикарибецу, Хоккайдо, Япония. J. Crust. Биол. 24: 496–501.
Никсират, Х., А. Куба и П. Козак. 2014. Морфологические изменения сперматозоидов и сперматофоров после спаривания
стенки раков Astacus leptodactylus: взгляд на неподвижный сперматозоид Anim.Репродукция. Sci. 149: 325–334.
Норо К.К., Д. да Силва-Кастильони, Л. Лопес-Греко, Л. Баккап и Г. Бонд-Бакап. 2007. Морфология семявыносящего протока
Parastacus defossus и P. varicosus и сравнение в пределах Parastacidae. Науплий 15: 43–48.
Noro, C.K., L.S. Лопес-Греко и Л. Баккап. 2008. Морфология гонад и тип сексуальности Parastacus defossus Faxon
1898, роющего интерсексуального рака из южной Бразилии (Decapoda: Parastacidae).Acta Zool. 89: 59–67.
Norrocky, M.J. 1991. Наблюдения за экологией, размножением и ростом роющих раков Fallicambarus
(Creaserinus) fodiens (Decapoda: Cambaridae) в северо-центральной части Огайо. Являюсь. Midl. Nat. 125: 75–86.
Паркин, С.М., К.Дж. Коллиер и Б.Дж.Хикс. 2002. Рост и динамика популяций раков Paranephrops planifrons в
ручьях в пределах естественных лесов и пастбищных угодий. Н.З. J. Mar. Freshwat. Res. 36: 847–861.
Парнес, С., И. Хайла, Г. Хулата и А. Саги. 2003. Определение пола у раков: являются ли интерсексуалы Cherax quadricarinatus (De cap od a,
Parastacidae) генетически самками? Genet. Res. 82: 107–116.
Pauly, D. and G. Gashutz. 1979. Простой метод согласования данных о колеблющемся росте длины с программой для карманных калькуляторов.
I.C.E.S.CM. 1979 / G: 24 Комитет по придонным рыбам, 26 стр.
Павлос Д., К. Формицки, А. Корзелецка-Оркиш и А. Винницки.2010. Процесс вылупления сигнального рака Pacifastacus
leniusculus (Dana, 1852) (Decapoda, Astacidae). Crustaceana 83: 1167–1180.
Пейн, Дж. Ф. 1972. История жизни Procambarus hayi. Являюсь. Midl. Nat. 87: 25–35.
Прайс, J.O. и Дж. Ф. Пэйн. 1984. Постэмбриональный рост и развитие раков Orconectes neglectus chaenodactylus
Williams, 1952 (Decapoda, Astacidea). Crustaceana 46: 176–194.
Рейнольдс, Дж. Д. 2002. Рост и воспроизводство.С. 152–191. В: D.M. Холдич (ред.). Биология пресноводных раков.
Blackwell Science Ltd., Лондон.
Рейнольдс, Дж. Д., Дж. Д. Селада, Дж. М. Каррал и М. А. Мэтьюз. 1992. Воспроизведение астцидных раков в неволе — текущие события
и их значение для культуры, с особым акцентом на Ирландию и Испанию. Invertebr. Репродукция. Dev. 22:
253–266.
Rhodes, C.P. и Д. Холдич. 1979. О размере и половом диморфизме Austropotamobius pallipes (Lereboullet): этап
в оценке потенциала коммерческой эксплуатации местных британских пресноводных раков.Аквакультура 17: 345–358.
Rode, A.L. and L.E. Бэбкок. 2003. Филогения ископаемых и существующих пресноводных раков и некоторых близкородственных нефропидов
омаров. Журнал биологии ракообразных 23: 418–435.
Rogowski, D.L., S. Sitko, S.A. Bonar. 2013. Оптимизация борьбы с инвазивными раками с использованием информации из истории их жизни.
Freshwat. Биол. 58: 1279–1291.
Руболини Д., П. Галеотти, Г. Феррари, М. Спайрани, Ф. Бернини и М. Фасола. 2006 г.Распределение сперматозоидов в зависимости от мужских признаков, размера самок
и поведения при совокуплении у пресноводных раков. Бех. Ecol. и Социобиол. 60: 212–219.
Руболини, Д., П. Галеотти, Ф. Пупин, Р. Сакки, П.А. Нарди и М. Фасола. 2007. Повторные спаривания и истощение сперматозоидов у пресноводных раков
Austropotamobius italicus. J. Freshwat. Биол. 52: 1898–1906.
Рудольф, Э. 1995a. Частичный протандрический гермафродитизм у роющего рака Parastacus nicoleti (Philippi, 1882)
(Decapoda: Parastacidae).J. Crust. Биол. 15: 720–732.
Рудольф, Э. 1995b. Случай гинандроморфизма у пресноводного рака Samastacus spinifrons (P hil ip pi, 188 2) (Dec ap oda,
Parastacidae). Crustaceana 68: 705–711.
Рудольф, Э. и К.С. Рохас. 2003. Эмбриональное и раннее постэмбриональное развитие роющих раков, Virila stac us
araucanius (Faxon, 1914) (Decapoda, Parastacidae) в лабораторных условиях. Ракообразные 76: 835–850.
Резерфорд, П.Л., Д. У. Данхэм и В. Эллисон. 1995. Победа в агонистических схватках самцов раков Orconectes rusticus
(Girard) (Decapoda, Cambaridae): размер чела имеет значение, а симметрия чела — нет. Crustaceana 68: 526–529.
Сент-Мари, Б. 2007. Спрос на сперму и распределение у десятиногих ракообразных. С. 191–210. В: Дж. Э. Даффи и М. Тиль (ред.).
Эволюционная экология социальных и половых систем. Издательство Оксфордского университета, Нью-Йорк.
Сэмми, Н. 1988. Биология размножения Cherax quadricarinatus на Северной территории.Материалы 1-й австралийской конференции по рыбным промыслам
, Технологический университет Кертина, Технологический университет Кертина: Перт, Вашингтон, Технологический университет Кертина
.
Scalici, M., A. Belluscio и G. Gibertini. 2008. Понимание структуры и динамики популяции раков, находящихся под угрозой исчезновения. J.
Zool. 275: 160–171.
Шольц, Г. 2002. Филогения и эволюция. С. 10–16. В: D.M. Холдич (ред.). Биология пресноводных раков. Блэквелл
Наука, Лондон.
Scholtz, G. and T. Kawai. 2002. Аспекты эмбрионального и постэмбрионального развития японских пресноводных раков
Cambaroides japonicus (Crustacea, Decapoda), включая гипотезу об эволюции материнской заботы в Astacida.
Acta Zoologica 83: 203–212.
Агонистические взаимодействия у раков Orconectes Quinebaugensis различаются по полу и сезону | Журнал биологии ракообразных
83″ data-legacy-id=»s1″> Введение
Агонистические взаимодействия были предметом интереса для многих таксонов, и формирование иерархии доминирования является особенно интригующим результатом этих взаимодействий во многих из этих случаев.Животные, которые участвуют в повторяющихся агонистических взаимодействиях в определенной области, образуют иерархию доминирования, которая позволяет контролировать ресурсы в популяции. Лица, получившие высокий социальный статус в иерархии доминирования, могут получить расширенный доступ к ресурсам. Например, было показано, что успешное агонистическое взаимодействие приводит к увеличению репродуктивного успеха у верветок ( Ceropithecus aethiop sabaeus ; Raleigh and McGuire, 1989) и тараканов ( Nauphoeta cinera ; Breed et al., 1980), а также льготный доступ к пищевым ресурсам радужной форели ( Oncorhynchus mykiss ; Johnsson, 1997) и пятнистых гиен ( Crocuta crocuta ; Tilson and Hamilton, 1984; обзор в Zulandt Schneider et al., 2001). Когда высокий социальный статус приводит к расширению доступа к ресурсам, это может привести к улучшению физической формы для лиц, которые доминируют в обществе (Moore, 2007). И наоборот, защита статуса доминирования в популяции может дорого обходиться индивидуальной физической форме, в основном за счет травм и / или энергии, затрачиваемой во время агонистических взаимодействий, а поддержание высокого статуса доминирования требует хорошего физического состояния для участия в повторяющихся агонистических взаимодействиях.Когда затраты на приспособленность и выгоды от участия в доминирующих взаимодействиях с сородичами потенциально велики, стратегии участия в этих взаимодействиях должны подвергаться строгому отбору, чтобы они максимизировали выгоды при одновременном снижении затрат (Maynard Smith, 1978). Кроме того, относительные затраты и выгоды от инвестирования в агонистические взаимодействия, вероятно, будут меняться на протяжении всей жизни человека, как с изменениями индивидуального состояния, так и с изменениями экологии и истории жизни.Например, потенциальные выгоды от высокого статуса доминирования могут варьироваться в зависимости от репродуктивного статуса людей в популяции или от наличия пищи или жилья.
Когда статус доминирования влияет на доступ индивида к партнерам, инвестиции в агонистические взаимодействия, скорее всего, находятся в рамках меж- и / или внутриполого отбора. Селективность самок в спаривании с доминирующими самцами была продемонстрирована у трехиглой колюшки ( Gasterosteus aculeatus ), где самки выборочно спариваются с самцами, яркая окраска которых отражает успех в агонистических столкновениях (Candolin, 1999), и у гуппи ( Poecilia reticulate ), где успех в агонистических столкновениях увеличивает репродуктивный успех, благодаря выбору самки, ранее непривлекательных самцов (Kodric-Brown, 1992).Было показано, что доминирующие самцы монополизируют имеющихся самок у европейского горчака ( Rhodeus sericeus ) независимо от предпочтений самок, которые впоследствии имеют ограниченный контроль над отцовством своего потомства (Reichard et al., 2005). В частности, когда существует сильная разница в инвестициях в потомство между мужчинами и женщинами, пол с более низкими инвестициями (обычно мужчины; Trivers, 1972) может получать относительно большие выгоды благодаря высокому социальному статусу.
У женщин также могут быть причины для инвестирования в доминирующие взаимодействия, которые, как и у мужчин, связаны с репродуктивными преимуществами, которые они могут получить.Однако, в то время как теория родительских инвестиций предсказывает, что самцы могут получить большие преимущества в фитнесе за счет многократного спаривания, женщины, по прогнозам, мало выиграют из-за полиандрии (Trivers, 1972), и, таким образом, польза для фитнеса от вложений в агонизм для раков может различаться по полу. Вложения женщин в агонизм вместо этого могут быть связаны с защитой потомства или ресурсов, необходимых потомству. Было показано, что наличие икры или молоди сопровождает повышенную агрессию самок раков (Figler et al., 1995b), и что эта тенденция к агрессивному поведению выше, чем у нематериальных женщин и у репродуктивно зрелых мужчин (Moore, 2007). В соответствии с гипотезой защиты потомства (Figler et al., 2001), такое поведение может привести к усилению защиты и, следовательно, к выживанию потомства в течение материнского сезона.
В этих условиях у самцов и самок, вероятно, будут развиваться совершенно разные стратегии инвестирования в агонистические взаимодействия с сородичами, и когда шанс спаривания ограничен во времени, это дифференциальное вложение должно быть таким же ограничено во времени.Например, самцы могут быть готовы вкладывать больше средств в агонизм, чем самки, особенно, когда вероятность спаривания высока, например, во время брачного сезона, но когда вероятность спаривания снижается во время периода без спаривания, самцы могут уменьшить свои шансы на спаривание. вложение в агонизм. Более того, для некоторых таксонов потенциальные выгоды от высокого социального статуса могут включать в себя множество ресурсов, таких как расширенный доступ как к пище, так и к партнерам, что приводит к динамической системе, в которой выгоды от агонизма меняются предсказуемо, например.g., сезонно или непредсказуемо, например, при изменении экологических условий.
Мы используем пресноводных раков, Orconectes quinebaugensis Mathews and Warren, 2008, чтобы проверить гипотезы об индивидуальных инвестициях в агонистические взаимодействия. Раки являются хорошими моделями для такого исследования, потому что, как и некоторые другие ракообразные, они структурированы иерархией доминирования, поддерживаемой визуальным отображением и боевыми действиями с участием крупных и потенциально опасных чела, а также посредством химиосенсорной коммуникации (Moore and Bergman, 2005).Кроме того, популяции раков умеренного пояса в Северной Америке структурированы в зависимости от сезонности изменений жизненного цикла (Hamr, 2002). Рост и линька раков происходит в основном летом, после чего самцы линяют и приобретают форму размножения I. Спаривание происходит осенью, и о полигамии у одного или обоих полов сообщалось у ряда видов (обзор у Galeotti et al., 2007), а у Orconectes placidus (Hagen, 1870) — множественное отцовство одиночных выводков. очевидно обычное явление (Walker et al., 2002). В течение зимы самки сохраняют сперму, которую они используют для оплодотворения яйцеклеток весной. Самки обеспечивают всю родительскую заботу о детенышах, включая выведение развивающихся эмбрионов и уход за только что вылупившимися детенышами в течение нескольких недель. Такой резко дифференцированный родительский вклад привел бы к сильному отбору самцов за конкуренцию за самцов через иерархию доминирования и, возможно, на самок за селективность спаривания.
Было показано, что на агрессию раков влияет множество факторов.Относительная важность определенных ресурсов, таких как еда или возможности для спаривания, может варьироваться в зависимости от ряда внутренних и внешних факторов, таких как пол (Figler et al., 2001), возраст (Schroeder and Huber, 2001), период размножения (Figler). et al., 2005), доступность ресурсов (Stocker and Huber, 2001) и наличие убежища (Bergman and Moore, 2003), так что конкуренция за ограниченные ресурсы может меняться во времени, и уровни агрессии могут соответственно меняться. Например, доминирующая особь может иметь возможность монополизировать пищевые ресурсы летом, когда рост имеет решающее значение для обоих полов при подготовке к осеннему сезону размножения.Монополизация доступного питания может привести к ускоренному росту человека и улучшению физической формы (Fero et al., 2007). Предыдущие исследования агрессии раков в основном были сосредоточены на механизмах установления доминирования, таких как роль химической коммуникации через мочу (обзор в Moore and Bergman, 2005), влияние предыдущего опыта на последующее поведение людей (Daws et al. , 2002; Bergman et al., 2003), эффекты состояния голода (Stocker and Huber, 2001) и компромисс между риском хищничества и возможностями спаривания (Pecor, 2006).Меньше внимания уделяется функции доминирования, такой как роль полового отбора в агонистическом поведении, связанном с доминированием (но см. Aquiloni et al., 2008; Fero et al., 2007). Здесь мы сообщаем о результатах наших экспериментов, чтобы понять, как особи O. quinebaugensis инвестируют в агонистические взаимодействия в качестве косвенного измерения инвестиций в доминирование. В частности, мы предсказали, что во время брачного сезона самцы (в размножающейся морфологии Формы I) будут вкладывать больше средств в внутриполовые конфликты, чем половозрелые самки.Кроме того, мы сравнили взаимодействия в период спаривания с взаимодействиями в период отсутствия спаривания, когда и самцы, и самки явно не репродуктивны. Мы предсказали, что не репродуктивные мужчины и женщины будут демонстрировать меньшую агрессию по сравнению с мужчинами формы I, и что не репродуктивные мужчины и женщины будут демонстрировать одинаковый уровень агрессии.
98″ data-legacy-id=»s2a»> Сбор и содержание животных
раков было собрано в реке Квинебауг в Стурбридже, Массачусетс, в августе – октябре 2007 г. для репродуктивной группы (всего 80) и в реках Квинебауг и Милл (в Блэкстоуне, Массачусетс) с апреля по июнь 2008 г. для нерепродуктивного сезона. (Всего 58 и 28 соответственно).В последнем случае из-за того, что раки линяют летом, мы не смогли получить достаточное количество особей между линьками на участке реки Квинебауг для адекватного размера выборки. У представителей этого вида брачный период примерно с октября по январь (личное наблюдение). В оба сезона репродуктивный статус самцов оценивался по морфологии гонопод; осенью все самцы были в Форме I, а летом все самцы были в Форме II. Репродуктивный статус самок оценивали по развитию желчных желез; осенью все самки имели полностью развитые голени, что указывало на половую зрелость; летом у всех самок не было развития гладких желез и не было высиживания эмбрионов или молоди.Кроме того, в осенних сборах все самки были собраны к началу сентября и помещены в лабораторию по отдельности, и, вероятно, не были спариваться. Таким образом, у нас было четыре лечебные группы (рис. 1). 2
Рис. 1
Среднее время, затраченное на физическое агонистическое поведение («захват») для каждой группы лечения, показано для больших и малых групп отдельно и вместе. Планки погрешностей представляют собой стандартную ошибку, а числа над каждой полосой указывают размеры выборки (каждая повторность представляет одну пару раков).
Рис. 1
Среднее время, затраченное на физическое агонистическое поведение («захват») для каждой группы лечения, показано для больших и малых групп отдельно и вместе. Планки погрешностей представляют собой стандартную ошибку, а числа над каждой полосой указывают размеры выборки (каждая повторность представляет одну пару раков).
Рис. 2
Доля испытаний, достигших максимального уровня интенсивности для групп после объединения. Цифры над каждой полосой указывают размеры выборки (каждая повторность представляет одну пару раков).Звездочки обозначают значимые отношения при | $ \ alpha = 0,0125 $ | .
Рис. 2
Доля испытаний, достигших максимального уровня интенсивности для групп после объединения. Цифры над каждой полосой указывают размеры выборки (каждая повторность представляет одну пару раков). Звездочки обозначают значимые отношения при | $ \ alpha = 0,0125 $ | .
Раки содержались в лаборатории Вустерского политехнического института (WPI) в закрытых рециркуляционных пресноводных системах с биологической, механической и УФ-фильтрацией.Все раки содержались не менее 2 недель перед использованием в испытании. Каждого раков помещали отдельно в | $ 4 \, {\ rm {L}} $ | пластиковый резервуар с глиняным горшком для укрытия, чтобы в период содержания особи находились в химическом контакте с другими раками, но никогда не контактировали физически. Раков кормили 3 раза в неделю на чередующейся диете из гранул коммерческих креветок и замороженной брокколи. Температура воды и световые условия имитировали естественные. Как для осенних, так и для летних сборов, раков были изолированы как физически, так и химически в течение одной недели перед использованием в испытаниях агрессии, поскольку было показано, что у некоторых видов химическая передача сигналов влияет на агонистическое поведение (Bergman and Moore, 2005).В течение этого времени каждое животное получало подмену воды на 50% каждый день, когда оно получало корм. Любая линька раков в течение 2 недель до или после использования в испытаниях была исключена из набора данных, как и любые испытания, в которых один противник умер в течение 6 дней после использования. Только одно испытание из большой репродуктивной мужской группы было исключено по прошествии отведенного времени.
05″ data-legacy-id=»s2c»> Сбор данных Парные противники были помещены в | $ 30,5 {\ text {cm width}} \ times 40.6 {\ text {cm length}} \ times 14.6 {\ text {cm depth}} $ | пластиковые емкости, наполненные свежей фильтрованной водопроводной водой; внутри танков противники были разделены прозрачными перегородками и получили возможность акклиматизироваться за | $ 2 \, {\ rm {min}} $ | . Затем разделители были сняты, и агонистические взаимодействия были записаны на видео для | $ 10 \, {\ rm {min}} $ | . По завершении каждого испытания обоих животных возвращали в их резервуары для хранения.Все резервуары тщательно промывались и наполнялись свежей фильтрованной водой между испытаниями. Видеозаписи испытаний оценивались слепым рецензентом за время, проведенное в физических агонистических взаимодействиях, обозначенных как «схватка» и определяемых как бокс, удары, хватание или толкание чела одним или обоими раками; и для проявления поведения, указывающего на различные уровни агрессивного взаимодействия (Таблица 1). Общее время, потраченное на «захватное» поведение, и доля схваток, достигших максимальной интенсивности, сравнивали между обработками как меры общего агонистического вложения.
Таблица 1 Этограмма окончательного поведения, используемая для приблизительного определения уровней интенсивности боя. Все виды поведения оценивались для обоих раков в паре. Этограмма адаптирована из Bruski & Dunham (1987) для использования в текущем исследовании.
Уровень интенсивности
. Описание
. 0 Нет взаимодействия 1 Приближение — приближение на одну длину тела противника, приводящее к отступлению противника в любом направлении 2 Выпад / угроза — быстрое приближение противника с Чела, поднятые на дисплее угрозы 3 Захват — бокс, удар, хватание или толкание чела одним или обоими раками 4 Переворот — раки повернуты на бок или спиной, когда их схватил с чела соперника
Уровень интенсивности
. Описание
. 0 Нет взаимодействия 1 Приближение — приближение на одну длину тела противника, приводящее к отступлению противника в любом направлении 2 Выпад / угроза — быстрое приближение противника с Челы, поднятые на дисплее угрозы 3 Захват — бокс, удар, хватание или толкание чела одним или обоими раками 4 Переворот — раки повернуты на бок или спиной, когда их схватил с чела оппонента
Таблица 1 Этограмма окончательного поведения, используемая для приблизительного определения уровней интенсивности боя.Все виды поведения оценивались для обоих раков в паре. Этограмма адаптирована из Bruski & Dunham (1987) для использования в текущем исследовании.
Уровень интенсивности
. Описание
. 0 Нет взаимодействия 1 Приближение — приближение на одну длину тела противника, приводящее к отступлению противника в любом направлении 2 Выпад / угроза — быстрое приближение противника с Чела, поднятые на дисплее угрозы 3 Захват — бокс, удар, хватание или толкание чела одним или обоими раками 4 Переворот — раки повернуты на бок или спиной, когда их схватил с чела соперника
Уровень интенсивности
. Описание
. 0 Без взаимодействия 1 Приближение — приближение на одну длину тела к противнику, приводящее к отступлению противника в любом направлении 2 Выпад / угроза — быстрое приближение противника с Челы, поднятые на дисплее угрозы 3 Захват — бокс, удар, хватание или толкание чела одним или обоими раками 4 Переворот — раки повернуты на бок или спиной, когда их схватил с чела оппонента
11″ data-legacy-id=»s3″> Результаты Не было значительной разницы в общем размере самок между падениями | $ (n = 34, \, \ bar {x} = 35.203 {\ text {mm}}) $ | и пружина | $ (n = 28, \, \ bar {x} = 34.993 {\ text {mm}}, \, t = 0.219, \, {\ text {DF}} = 60, \, P = 0.827 ) $ | , либо для женщин в маленьком | $ ({\ text {fall:}} = 11, \, \ bar {x} = 30.572 {\ text {mm}} $ | ; | $ {\ текст {весна:}} n = 11, \, \ bar {x} = 31,046 {\ text {mm}}, \, t = -0,555, \, {\ text {DF}} = 20, \, P = 0,585 $ | ) или большой | $ ({\ text {fall:}} n = 23, \, \ bar {x} = 37,417 {\ text {mm}}; \, {\ text {spring:}} n = 17, \, \ bar {x} = 37,547 {\ text {mm}}, \, t = — 0,214, \, {\ text {DF}} \, = 38, \, P = 0.832) $ | размерные классы. Наблюдалась значительная разница в общем размере мужчин между падениями | $ (n = 34, \, \ bar {x} = 35.247 {\ text {mm}}) $ | и пружина | $ (n = 42, \, \ bar {x} = 33,136 {\ text {mm}}, \, t = 2,867, \, {\ text {DF}} = 74, \, P = 0,005 ) $ | , но не для маленьких | $ ({\ rm {fall}}: \, \, n = {\ rm {14}}, \, \, \ bar x = {\ rm {32} } .000 \, \, {\ rm {mm}}; \, \, {\ rm {spring}}: \, \, n = {\ rm {24}}, \, \, \ bar x = { \ rm {3}} 0. {\ rm {746}} \, \, {\ rm {mm}}, \, \, t = {\ rm {\ bar 2}}. {\ rm {376}} , \, \, {\ rm {DF}} = {\ rm {36}}, \, \, P = 0.0 {\ rm {23}}) $ | или большой | $ ({\ text {fall:}} n = 20, \, \ bar {x} = 37,520 {\ text {mm}}; \, {\ text {spring:}} n = 18, \ , \ bar {x} = 36,322 {\ text {mm}}, \, t = 2,275, \, {\ text {DF}} = 36, \, P = 0,029) $ | группы отдельно.
При сравнении времени, затраченного на общее агонистическое поведение, и лечение, и размер показали значительный эффект в различных сравнениях (таблица 2, рис. 1). Самцы | $ (n = 17) $ | и женщины | $ (n = 22) $ | в осеннем репродуктивном сезоне было обнаружено значительное влияние секса на общее время, затрачиваемое на агонистическое поведение, при этом самцы тратят больше времени на такое поведение, чем самки, и отсутствие значительного влияния размера на общее агонистическое поведение в этих двух группах.Существенной разницы между мужчинами | $ (n = 21) $ | и женщины | $ (n = 19) $ | летом, в период отсутствия репродукции, общее время, проведенное в агонистическом поведении, но и самцы, и самки для этого сравнения показали значительный эффект размера, при этом более крупные животные тратят значительно больше времени на агонистическое поведение, чем более мелкие животные. Сравнение репродуктивных и не репродуктивных особей не показало значительных различий в общем времени, проведенном в агонистическом поведении в зависимости от сезона или размера для любого пола, хотя для женщин взаимодействие между репродуктивным состоянием и размером имело значительный эффект и сохранялось в модель (но которая не имела значения при исправлении Бонферрони | $ {\ rm {\ alpha}} = 0.0125 $ | ).
Таблица 2 Результаты дисперсионного анализа с фиксированными эффектами сезона, например, репродуктивные самцы по сравнению с непродуктивными самцами, или пол, например, репродуктивные самцы по сравнению с репродуктивными самками. Во всех сравнениях размер был включен как фактор; член взаимодействия был незначительным | $ (P> 0,05) $ | во всех случаях, кроме сравнения репродуктивных самок с непродуктивными самками, и был исключен из анализов, в которых он не был значимым.
. Фактор 1 (размер)
. Фактор 2 (сезон или пол)
. Фактор 1 * фактор2
. Контрастность
. ф
. | $ P $ |
. ф
. | $ P $ |
. ф
. | $ P $ |
. Между сезонами: 1.Репродуктивные самцы и непродуктивные самцы 0,468 0,498 1,174 0,286 — — 2. Репродуктивные самки и непродуктивные самки 4,820 0,034 0,146 4,027 0,052 Между полами: 3.Репродуктивные самцы и репродуктивные самки 0,065 0,801 9,071 0,005 * — — 4. Непродуктивные самцы и непродуктивные самки 7,381 9 * 0,025 0,876 — —
. Фактор 1 (размер)
. Фактор 2 (сезон или пол)
. Фактор 1 * фактор2
. Контрастность
. ф
. | $ P $ |
. ф
. | $ P $ |
. ф
. | $ P $ |
. Между сезонами: 1.Репродуктивные самцы и непродуктивные самцы 0,468 0,498 1,174 0,286 — — 2. Репродуктивные самки и непродуктивные самки 4,820 0,034 0,146 4,027 0,052 Между полами: 3.Репродуктивные самцы и репродуктивные самки 0,065 0,801 9,071 0,005 * — — 4. Непродуктивные самцы и непродуктивные самки 7,381 9 * 0,025 0,876 — —
Таблица 2 Результаты ANOVA с фиксированными эффектами сезона, например, репродуктивные самцы по сравнению с непродуктивными самцами или полом, e.g, репродуктивные самцы по сравнению с репродуктивными самками. Во всех сравнениях размер был включен как фактор; член взаимодействия был незначительным | $ (P> 0,05) $ | во всех случаях, кроме сравнения репродуктивных самок с непродуктивными самками, и был исключен из анализов, в которых он не был значимым.
. Фактор 1 (размер)
. Фактор 2 (сезон или пол)
. Фактор 1 * фактор2
. Контрастность
. ф
. | $ P $ |
. ф
. | $ P $ |
. ф
. | $ P $ |
. Между сезонами: 1. Репродуктивные самцы по сравнению с непродуктивными самцами 0.468 0,498 1,174 0,286 — — 2. Репродуктивные самки по сравнению с непродуктивными самками 4,820 0,034 2,207 0,146 4,0 Между полами: 3. Репродуктивные самцы и репродуктивные самки 0.065 0,801 9,071 0,005 * — — 4. Непродуктивные самцы и непродуктивные самки 7,381 0,010 * 76 — —
. Фактор 1 (размер)
. Фактор 2 (сезон или пол)
. Фактор 1 * фактор2
. Контрастность
. ф
. | $ P $ |
. ф
. | $ P $ |
. ф
. | $ P $ |
. Между сезонами: 1.Репродуктивные самцы и непродуктивные самцы 0,468 0,498 1,174 0,286 — — 2. Репродуктивные самки и непродуктивные самки 4,820 0,034 0,146 4,027 0,052 Между полами: 3.Репродуктивные самцы и репродуктивные самки 0,065 0,801 9,071 0,005 * — — 4. Непродуктивные самцы и непродуктивные самки 7,381 9 * 0,025 0,876 — —
Мы обнаружили аналогичные эффекты размера и лечения, когда сравнивали долю драк, достигших максимального уровня интенсивности.Во всех случаях, кроме одного (самки летом, нерепродуктивный сезон, | $ P = 0,018 $ | ), не было существенной разницы в доле испытаний, достигающих максимальной интенсивности между группами малого и большого размера (репродуктивные самцы | $ P = 0,153 $ | , репродуктивные самки | $ P = 0,364 $ | , непродуктивные самцы | $ P = 0,367 $ | ). Следовательно, для трех последних курсов лечения были объединены большие и малые группы. Как и в случае продолжительности агонистических взаимодействий, значительно больше драк между репродуктивными самцами достигли максимальной интенсивности, чем между репродуктивными самками | $ (P = 0.001) $ | . Не было обнаружено существенной разницы в доле, достигающей максимальной интенсивности между непродуктивными самцами и крупными не репродуктивными самками летом | $ (P = 0,721) $ | ; тем не менее, значительно больше драк между не репродуктивными самцами достигло максимальной интенсивности, чем между маленькими не репродуктивными самками | $ (P = 0,003) $ | . Точно так же значительно больше испытаний между крупными непродуктивными самками достигло максимальной интенсивности, чем испытания между репродуктивными самками | $ (P = 0,020) $ |, в то время как не было значительной разницы в интенсивности между маленькими непродуктивными самками и репродуктивными самками | $ ( Р = 1.000) $ | . Не было существенной разницы в доле испытаний, достигающих максимальной интенсивности между самцами в репродуктивном и не репродуктивном периодах | $ (P = 0,743) $ | .
Обсуждение
Существует множество факторов, как внутренних (размер тела и чела, пол, репродуктивное состояние, предыдущий социальный опыт, мотивационное состояние, нейрохимия), так и внешних (факторы общения с окружающей средой, воспринимаемая ценность ресурсов), которые влияют на количество времени и риски для индивидов. готовы вкладывать средства в агонистические взаимодействия (Мур, 2007).Наши данные показывают, что самцы O. quinebaugensis проводили значительно больше времени в агонистическом поведении и участвовали в большей части драк с максимальной интенсивностью, чем самки в осеннем репродуктивном сезоне, как мы и предсказывали. Это может указывать на то, что самцы реагируют на половой отбор, вкладывая больше энергии в агонизм, если доминирующие самцы имеют репродуктивное преимущество.
Если доминирование действительно связано с повышенной приспособленностью самцов, можно ожидать, что в течение репродуктивного сезона, когда этот ресурс является наиболее ценным, будет инвестировано больше времени и, следовательно, больший риск.Другие исследования показали, что интенсивность и продолжительность агонистических встреч коррелируют с ценностью ресурса и его доступностью для участников (Bergman and Moore, 2003; Stocker and Huber, 2001; обзор в Moore, 2007). У раков, например, материнские самки (несущие икру или молодые особи) могут быть намного более агрессивными, чем нематериальные самки (Figler et al., 1995b), весной, когда качество и выживаемость молоди напрямую влияют на приспособленность самок. В нашем эксперименте, однако, не было значительной разницы во времени, затраченном на агонизм, или пропорции схваток, достигающих максимальной интенсивности, между репродуктивными и не репродуктивными самцами.В частности, в классах большого размера для анализа продолжительности самцы Формы II проводили столько же времени в агрессии, как и самцы Формы I. Точно так же крупные непродуктивные самки проводили больше времени в агрессии и имели более высокую долю драк с максимальной интенсивностью, чем репродуктивные самки. Хотя эта разница была значимой в отношении интенсивности борьбы, она не была значимой для времени, проведенного в агонистическом поведении, вероятно, в результате сильного эффекта размера у не репродуктивных самок.В целом, в то время как поведение репродуктивных самцов и самок согласуется с нашей гипотезой о том, что половой отбор стимулирует инвестиции в агонизм, данные по нерепродуктивным ракам — нет. Мы предлагаем два альтернативных объяснения этого наблюдения.
Во-первых, летний сезон представляет собой основной период роста как молоди, так и взрослых раков O. quinebaugensis ; такие сезонные закономерности линьки и роста обычны для раков умеренного климата (Reynolds, 2002).В этих условиях ценность таких ресурсов, как еда и жилье, может возрасти как для мужчин, так и для женщин. Монополизация пищевых ресурсов может привести к ускоренному росту и улучшению индивидуальной физической формы (Fero et al., 2007), а размер часто положительно коррелирует с результатами борьбы раков (например, Pavey and Fielder, 1996; Daws et al., 2002). ; Schroeder, Huber, 2001; Figler et al., 1995a; Klocker and Strayer, 2004; обзор в Moore, 2007) и плодовитость у самок (Rubolini et al., 2006; Reynolds, 2002), что потенциально может привести к репродуктивным преимуществам на более позднем этапе жизненного цикла для обоих полов. Укрытия также могут быть ресурсом равной ценности, который потребовал бы увеличения агонистических вложений для обоих полов, поскольку частая линька также увеличивает долю времени, в течение которого рак становится более уязвимым для хищников (Hamr, 2002). Следовательно, летом мужчины и женщины могут одинаково вкладывать средства в агонизм, потому что наиболее ценные ресурсы одинаково важны для обоих полов. Осенью относительная важность ресурсов может измениться в ответ на замедление роста и охлаждение температуры воды, а инвестиции в конкуренцию за партнеров, а не за еду или жилье отражаются в ориентированных на пол вложения в агонизм.
В качестве альтернативы, самки во время репродуктивного сезона могут извлечь большую пользу из воздержания от этих увеличенных агонистических вложений в пользу других энергетических потребностей. Для мужчин было высказано предположение, что инвестиции в агонистические взаимодействия должны конкурировать с инвестициями в другие компоненты, связанные с приспособленностью, такие как привлечение партнера или родительская забота (Qvarnström, 1997; Griffith and Sheldon, 2001), и можно было бы ожидать аналогичной торговли. снижение расхода энергии для женщин (например,г., Ветцель, 2002; Бернардо, 1996). Первичная репродуктивная деятельность самок в это время заключается в формировании яиц (Galeotti et al., 2006), что, вероятно, требует больших затрат энергии. Самки могут получить больше преимуществ в фитнесе, вкладывая больше средств в производство яиц, а не борясь за доступ к партнерам или пище, поскольку репродуктивный успех самок ограничен количеством яиц, которые она производит, а также качеством и выживаемостью ее потомства (Trivers, 1972). Было высказано предположение, что размер яйца является решающим фактором, определяющим приспособленность потомства (Bernardo, 1996), а вложение энергии в схватку с аналогами может ограничить способность самки производить большие яйца или кладку.
Поиск партнера и дискриминация также могут повлечь за собой энергетические затраты для женщин (Kokko et al., 2002). Инвестиции в агонизм в репродуктивном сезоне также могут отвлекать от времени и энергии, которые самки могут тратить на отбор и спаривание желаемых, высококачественных самцов, что в конечном итоге может повлиять на качество их потомства (Kokko et al., 2002; Berglund et al. ., 1996). У таких видов, как O. quinebaugensis , у которых самки хранят сперму и откладывают оплодотворение до нескольких месяцев после спаривания (Hamr, 2002), самки также могут получить косвенные выгоды от выделения времени и энергии на получение нескольких спариваний (Jennions and Petrie, 2000).Любые травмы, полученные в результате агрессивного поведения во время репродуктивного сезона, также могут помешать самкам спариваться с желаемым самцом или вообще успешно спариваться.
Весной, когда самка вкладывает средства в потомство в виде эмбрионов и частично независимых молодых особей, самки могут извлечь выгоду за счет повышенной агрессии, если это позволяет им оставаться в приюте, предположительно как компонент защиты потомства (Фиглер). и другие., 2001). Мур (2007) в своем обзоре соответствующей литературы обнаружил, что самки матери были более агрессивными, чем самки нематериального происхождения и репродуктивные самцы, а Фиглер и др. (2001) предположили, что это было частично связано с внутренним состоянием, поскольку даже самки матери с удаленным потомством показали более высокую защиту укрытия по сравнению с самками нематериального происхождения при сравнении предыдущих исследований. В совокупности это говорит о том, что вклад женщин в агонистические взаимодействия может происходить в рамках некоторой формы полового отбора, но в другом направлении, чем можно было бы ожидать от мужчин.
Мы также сообщаем противоречивые данные о влиянии размера на агонистическое поведение. Летом наблюдалось значительное влияние размерной группы как для самцов, так и для самок в зависимости от продолжительности: более крупные раки обоих полов проводили больше времени в агонистическом поведении, чем более мелкие раки. Также наблюдалось влияние размера на интенсивность агрессии между самками летом: у крупных самок была более высокая доля драк, достигающих максимальной интенсивности, чем у маленьких самок. Принято считать, что размер является основным фактором, который обычно предсказывает исход и агрессивное состояние при взаимодействии раков (Pavey and Fielder, 1996; Daws et al., 2002; Клокер и Страйер, 2004; Figler et al., 1995a; Шредер и Хубер, 2001; рассмотрено в Moore, 2007), и было высказано предположение, что маленькие и большие раки могут применять разные стратегии борьбы в зависимости от ценности ресурсов и риска для человека (Schroeder and Huber, 2001). Более крупные, более доминирующие люди потенциально меньше рискуют вступить в агонистическое поведение, поскольку более вероятно, что они добьются успеха, особенно в паре с меньшим противником. Однако мы не наблюдали подобных эффектов размера в брачный период, когда и большие, и маленькие особи обоих полов одинаково вкладывали средства в драки.
Хотя объяснение этому наблюдению неясно, мы отмечаем, что в нерепродуктивной коллекции животных некоторые (возможно, многие) особи обоих полов в группе меньшего размера, возможно, никогда не претерпели периода воспроизводства, в то время как особи в группах большего размера вероятность воспроизводства выше в предыдущем году. Этим можно объяснить значительную разницу, наблюдаемую в доле схваток, достигающих максимальной интенсивности, между репродуктивными самками осенью и крупными непродуктивными самками летом, в то время как сравнение репродуктивных самок осенью и мелких непродуктивных самок летом было незначительным.Материнские самки других видов раков проявляют повышенную агрессию по отношению как к репродуктивным самцам, так и к нематеринским самкам (Figler et al, 1995b), и предполагается, что это частично связано с изменением внутреннего состояния (Figler et al. ., 2001). Следовательно, возможно, что у крупных самок летом мог быть некоторый остаточный внутренний эффект, связанный с недавно рассеяной кладкой молоди, тогда как у маленьких самок меньше шансов иметь недавнюю кладку, хотя недавнее материнское состояние большинства самок в летняя группа была неизвестна.
В целом, наши данные показывают, что индивидуальные вложения в агонизм особей O. quinebaugensis различаются в зависимости от сезона, пола и размера. Тем не менее, мы специально выдвинули гипотезу о том, что взаимодействия с доминированием являются сексуально отобранным поведением, и предсказали, что репродуктивные самцы будут проявлять повышенную агрессию по сравнению как с репродуктивными самками, так и с непродуктивными раками, и наши данные не обеспечивают убедительной поддержки этой гипотезы. Вместо этого наши данные показывают, что агонистические взаимодействия у O.quinebaugensis может регулироваться сложным и динамичным набором индивидуальных затрат и выгод, и это дополнительное экспериментирование может помочь выяснить роль окружающей среды, жизненного цикла и других факторов в социальном поведении этого таксона.
Благодарности
Мы хотели бы поблагодарить Э. Райдер за ее помощь в анализе данных и следующих студентов за их помощь в сборе данных: Л. Амендола, Э. Гринуэй, К. Рикер, Р.Стэнфорд, У. Терротт, Т. Малхерн и М. Беккер. Наконец, мы хотели бы поблагодарить Инженерный корпус армии США за предоставление разрешения на специальное использование (Л. М.) для сбора раков на плотине Ист-Бримфилд в Стербридже, Массачусетс.
Список литературы
Berglund
A.
, Bisazza
A.
и Pilastro
A.
1996
Вооружения и украшения: эволюционное объяснение свойств двойной полезности.
Биологический журнал Линнейского общества
58
385
— 399
Бергман
DA
и Мур
PA
2003
Полевые наблюдения за внутривидовым агонистическим поведением двух видов раков Orconectes rusticus и Orconectes virilis , в разных местообитаниях.
Биологический бюллетень
205
26
— 35
Бергман
D.A.
и Moore
P. A.
2005
Продолжительное воздействие социальных запахов изменяет последующие социальные взаимодействия у раков Orconectes rusticus .
Поведение животных
70
311
— 318
Бергман
DA
, Козловски
К.
, Макинтайр
JC
, Хубер
, Р.
, А.G.
и Moore
P. A.
2003
Временная динамика и связь эффектов победителя у раков, Orconectes rusticus .
Поведение
140
805
— 825
Бернардо
J.
1996
Особый материнский эффект размера отростка, особенно размера яйца: модели, модели, качество доказательств и интерпретаций.
Американский зоолог
36
216
— 236
Порода
M.D.
, Smith
S. K.
и Gall
B. G.
1980
Поведенческие стратегии во время агонистических взаимодействий между самцами у тараканов.
Поведение животных
28
1063
— 1069
Кандолин
U.
1999
Соревнования самцов и самцов облегчают выбор самок колюшки.
Труды Лондонского королевского общества B: Биологические науки
266
785
— 789
Кокс
C.R.
и Le Boeuf
B. J.
1977
Подстрекательство самок к мужской конкуренции: механизм полового отбора.
Американский натуралист
111
317
— 335
Daws
AG
, Грили
J.
, Konzen
K.
и Moore
PA
2002
Предыдущий опыт изменяет исход агрессивных взаимодействий между самцами раков, Procambarus clarkii .
Поведение и физиология в морской и пресноводной среде
35
139
— 148
Fero
K.
, Simon
JS
, Jourdie
V.
и Moore
Moore
2007
Последствия социального доминирования для использования ресурсов раков.
Поведение
144
61
— 82
Фиглер
M.H.
, Finkelstein
JE
, Twum
M.
и Peeke
HVS
1995a
Самцы красных болотных раков, Procambarus clarkii , сразу преобладают более мелкие представители традиционных сообществ. , смешанный сородич.
Агрессивное поведение
21
225
— 236
Фиглер
M. H.
, Twum
M.
, Finkelstein
JE
и Peeke
HVS
1995b
Материнская агрессия у красных болотных раков ( Procambarus clarkii , Girard): связь между репродуктивным статусом и исходом агрессивных встреч с самцами и самками сородичи.
Поведение
132
107
— 125
Фиглер
M. H.
, Blank
G. S.
и Peeke
H. V. S.
2001
Материнская территориальность как стратегия защиты потомства у красных болотных раков Procambarus clarkii , Girard.
Агрессивное поведение
27
391
— 403
Фиглер
MH
, Blank
GS
и Peeke
HVS
2005
HVS
2005
Resident Shelter Procambarus clarkii Жирар и другие злоумышленники, различающиеся по полу и репродуктивному статусу.
Поведение и физиология в морской и пресноводной среде
38
237
— 248
Galeotti
P.
, Pupin
F.
, Rubolini
D.
, Sacchi
, Sacchi
, Nardi
PA
и Fasola
M.
2007
Влияние статуса спаривания на поведение при копуляции и расход спермы у пресноводных раков Austropotamobius italicus .
Поведенческая экология и социобиология
61
711
— 718
Галеотти
P.
, Rubolini
D.
, Fea
G.
, Ghia
Nardi
P.
, Gherardi
F.
и Fasola
M.
2006
Самки пресноводных раков корректируют размер яиц и кладок в зависимости от нескольких признаков самцов.
Труды Лондонского королевского общества (B)
273
1105
— 1110
Griffith
S.
и Sheldon
B.
2001
Фенотипическая пластичность в выражении сексуально отобранных черт: игнорируемые компоненты вариации.
Поведение животных
61
987
— 993
Guiasu
R. C.
и Dunham
D.W.
1998
Межформные агонистические соревнования у самцов раков, Cambarus robustus (Decapoda, Cambaridae).
Биология беспозвоночных
117
144
— 154
Hagen
HA
1870
Иллюстрированный каталог Музея сравнительной зоологии Гарвардского колледжа
2
11 viii + 109 страниц
Hamr
P.
2002
Orconectes .
585
— 608
Holdich
DM
Биология пресноводных раков
Blackwell Science
Oxford
Hays
WL
1994
Statistics
Harcourt
Jennions
M.
и Petrie
M.
2000
Почему самки размножаются? Обзор генетических преимуществ.
Биологические обзоры
75
21
— 64
Johnsson
J. I.
1997
Индивидуальное распознавание влияет на агрессию и отношения доминирования у радужной форели, Oncorhynchus mykiss .
Этология
103
267
— 282
Кирк
RE
1995
Экспериментальный план: процедуры для поведенческих наук
Brooks / Cole Publishing Company
2 Pacific Grove4,
, Pacific Grove4,
,
, Pacific Grove4,
Клокер
С.A.
и Strayer
D. L.
2004
Взаимодействия среди инвазивных раков ( Orconectes rusticus ), местных раков ( Orconectes limosus ) и местных двустворчатых моллюсков (Sphaeriidae и Unionidae).
Северо-восточный натуралист
11
167
— 178
Кодрик-Браун
A.
1992
Доминирование самцов может повысить успешность спаривания у гуппи.
Поведение животных
44
165
— 167
Кокко
H.
, Brooks
R.
, Jennions
M.
и Morley
J.
2002 Эволюция выбора партнера и предубеждений в отношении спаривания.
Труды Лондонского королевского общества (B)
270
653
— 664
Мэтьюз
Л. М.
и Warren
A. H.
2008
Новый рак из рода Orconectes Cope, 1872 г. из южной части Новой Англии (Crustacea: Decapoda: Cambariidae).
Труды Вашингтонского биологического общества
121
374
— 381
Мейнард Смит
Дж.
1978
Теория оптимизации в эволюции.
Ежегодный обзор экологии и систематики
9
31
— 56
Мур
P.A.
2007
Агонистическое поведение пресноводных раков — влияние внутренних и внешних факторов на агрессивные встречи и доминирование.
90
— 114
Даффи
EJ
и Тиль
M.
Эволюционная экология социальных и половых систем — ракообразные как модельные организмы
Harvard University Press
New York
Moore
PA
и Bergman
D.A.
2005
Запах успеха и неудачи: роль внутренних и внешних химических сигналов в социальном поведении раков.
Интегративная и сравнительная биология
45
650
— 657
Pavey
CR
и Fielder
DR
1996
Влияние разницы в размерах на агонистическое поведение пресноводных раков Cherax cuspidatas (Decapoda: Parastacidae).
Journal of Zoology
238
445
— 457
Pecor
K. W.
2006
Компромисс между преследованием и предотвращением хищничества у зрелых раков, Orconectes virilis .
Поведение и физиология в морской и пресноводной среде
39
229
— 233
Qvarnstrom
A.
1997
Экспериментально увеличенный размер значка увеличивает конкуренцию мужчин и снижает заботу родителей о самцах в ошейниковой мухоловке.
Протоколы Лондонского королевского общества (B)
264
1225
— 1231
Qvarnstrom
A.
и Forsgren
E.
1998
Если женщины предпочитают доминирующих мужчин ?
Тенденции в экологии и эволюции
13
498
— 501
Роли
MJ
и McGuire
MT
1989
Влияние самок на доминирование самцов у 9000 обезьян в неволе7 Cercopithecus aethiops sabaeus .
Поведение животных
38
59
— 67
Reichard
M.
, Bryja
J.
, Ondračková
M.
, Dávidová
, P.
и Smith
C.
2005
Половой отбор на доминирование самца снижает возможности выбора самки у европейского горчака ( Rhodeus sericeus ).
Молекулярная экология
14
1533
— 1542
Рейнольдс
Дж. Д.
2002
Рост и воспроизводство.
152
— 191
Holdich
DM
Биология пресноводных раков
Blackwell Science
Oxford
Rubolini
D.
, Galeotti
P.
, Ferrari
, Спайрани
М.
, Bernini
F.
и Fasola
M.
2006
Распределение сперматозоидов в зависимости от мужских признаков, размера самки и поведения при совокуплении у пресноводных раков.
Поведенческая экология и социобиология
60
212
— 219
Schroeder
L.
и Huber
R.
2001
Стратегии борьбы различаются размером и аллометрическим ростом раки, Orconectes rusticus .
Поведение
138
1437
— 1449
Stocker
AM
и Huber
R.
2001
Стратегии борьбы с раками Orconectes rustidae состояние голода и наличие пищевых сигналов.
Этология
107
727
— 736
Tilson
R.H.
и Hamilton
W.J.
III 1984
Социальное доминирование и особенности питания пятнистых гиен.
Поведение животных
32
715
— 724
Trivers
R. L.
1972
Родительское вложение и половой отбор.
139
— 179
Campbell
B.
Половой отбор и происхождение человека
Aldine
Chicago
Walker
D.
, Porter
B.A.
и Avise
J. C.
2002
Оценка генетического родства у раков Orconectes placidus , беспозвоночное с высокой плодовитостью, нуждающееся в расширенном материнском уходе.
Молекулярная экология
11
2115
— 2122
Wetzel
J.
2002
Изменение формы взрослых самок раков рода Orconectes (Decapoda: Cambaridae).
American Midland Naturalist
147
326
— 337
Winer
BJ
, Brown
DR
и Michels
KM
1991
Принципы статистического проектирования
McGraw-Hill
New York
Zulandt-Schneider
RA
, Huber
R.
и Moore
P.A.
2001
Распознавание индивидуума и статуса у раков, Orconectes rusticus : влияние выделения мочи на динамику схватки.
Поведение
138
137
— 153
© The Crustacean Society, 2016. Опубликовано Brill NV, Leiden
Раки-мутанты, полностью самки, размножающиеся путем клонирования
Марморреба сейчас настолько распространены на Мадагаскаре, что они угрожают местным видам.Андре Карват / CC BY-SA 2.5 Мраморные раки очень похожи на любых других пресноводных ракообразных. У него два когтя, десять ног и красивый сине-коричневый мраморный панцирь. Однако это шестидюймовое существо, обитающее в ручьях и озерах по всему миру, представляет гораздо более зловещие возможности, чем вы могли ожидать. Его новое научное название дает несколько подсказок: Procambarus virginalis. Все мраморные раки — самки, и они размножаются путем клонирования самих себя.
Фрэнк Лыко, биолог из Немецкого центра исследования рака, впервые услышал о мраморных раках от владельца аквариума, который в 1995 году подобрал «техасских раков» в зоомагазине.Они были поразительно большими и откладывали огромные партии яиц — сотни за один присест. Вскоре, как сообщает New York Times , любитель был осажден таким количеством раков, что раздавал их своим друзьям. И вскоре после этого marmorkrebs, как их называют по-немецки, стали появляться в зоомагазинах в Европе.
В этих раках было что-то очень странное. Все они были самками, и все они отложили сотни яиц без спаривания. Из этих яиц, в свою очередь, вылупились еще сотни самок, каждая из которых вырастает полностью способной к воспроизводству.В 2003 году ученые секвенировали их ДНК и подтвердили то, что многие владельцы уже считали так: каждый детеныш раков был клоном своей матери, и они наполняли рыбные ловушки Европы с угрожающей скоростью. «Люди начинают с одного животного, а через год у них будет пара сотен», — сказал Лико в интервью Times .
Всего 25 лет назад мраморных раков вообще не существовало. Теперь их можно найти в дикой природе миллионами в Германии, Чехии, Венгрии, Хорватии, Украине, Японии и Мадагаскаре.
Онлайн-совет для владельцев марморкребов ясен: они размножаются, как кролики. «Хотя 10 галлонов [аквариума] хватит на пару месяцев, — предупреждает один сайт, — эти 10 галлонов будут становиться все меньше и меньше по мере размножения этих животных. … Хорошее практическое правило: чем больше колония, тем больше аквариум. Начните с 40 галлонов и увеличьте объем, если можете себе это позволить ». Любители открыли эту истину на свой счет, что побудило многих выбрасывать своих дополнительных домашних животных в озера и ручьи.
В любых условиях раки прекрасно себя чувствуют.Оказывается, Marmorkrebs настолько же выносливы, насколько и плодовиты. А когда в водоеме их слишком много, они просто перемещаются. Times сообщает, что они пройдут сотни ярдов, чтобы найти новые озера или ручьи, где один экземпляр может произвести целую популяцию.
За последние пять лет Лико и его коллеги секвенировали геном мраморных раков, опубликовав свои результаты на этой неделе в журнале Nature Ecology & Evolution .Похоже, что эти раки являются результатом единственной радикальной мутации у речного рака, обитающего в реке Сатилла во Флориде и Джорджии. Около трех десятилетий назад спарились два плоских рака. При нормальном воспроизводстве каждая половая клетка имеет по одной копии каждой хромосомы. Но с одним из этих двух первых раков было что-то не так — мутация, которая оставила у него две копии в его половой клетке. Почему-то потомство оказалось выносливым и в отличном состоянии. Невероятно, но у него было по три копии каждой хромосомы и способность к бесполому размножению, не требуя участия мужчины.
Итак, захватит ли Землю армия клонов самок мраморных раков? «Это зависит от обстоятельств», — сказал Лыко « Times ». «Может быть, они просто проживут 100 000 лет. Лично для меня это займет много времени, но в процессе эволюции это будет просто отметкой на радаре ». По его словам, у их бесполого размножения есть свои преимущества и недостатки. Рост популяции стремительный, один экземпляр способен произвести сотни плодовитых потомков. Но они тоже уязвимы. При отсутствии генетических вариаций, если болезнь может уничтожить один клон, все остальные с большой вероятностью будут уничтожены.Однако сколько времени потребуется, чтобы появился такой патоген, остается загадкой. А пока во всем мире раки чувствуют себя как дома.
ADW: Cambarus bartonii: ИНФОРМАЦИЯ
Географический диапазон
Cambarus bartonii обитает в восточной и южной частях Соединенных Штатов, а также в юго-восточной части Канады. Однако чаще всего они встречаются в диапазоне от Нью-Брансуика, Канада, до северной Джорджии и в восточных частях Кентукки и Теннесси.(Зеленый, 1967)
Среда обитания
Cambarus bartonii обитают на дне ручьев, ручьев, малых рек и озер. Они строят норы, иногда называемые «дымоходами». Их норы могут быть простыми дуплами под камнем или более замысловатыми, с боковыми проходами. Дымоходы расположены вдоль кромки воды. Большая часть конструкции находится под водой, но вершина выступает и напоминает дымоход. Дымоходы различаются по размеру, самое большое отверстие составляет около восьми сантиметров.(Макман, 1960)
- озера и пруды
- реки и ручьи
Физическое описание
Cambarus bartonii — пресноводное ракообразное. При первом осмотре он выглядит «как омар». У него острая морда, а глаза на подвижных стеблях. Тонкий, но прочный экзоскелет темно-коричневого цвета, иногда со слегка красным оттенком.Точный оттенок зависит от того, как выглядит нижний субстрат среды обитания.
Из придатков наиболее заметны хелипеды. Эти придатки прикрепляются к грудной клетке и также называются первыми ходовыми ногами. Однако у этих придатков есть когти, и они используются для защиты и ловли пищи, а не для ходьбы. Cambarus bartonii также имеет четыре другие пары ходильных ног, прикрепленных к грудной клетке. Брюшные придатки Cambarus bartonii называются пловцами или плеоподами, они намного меньше ходячих ног и не подходят для плавания.Мужские пловцы модифицированы для передачи пакетов спермы самкам во время размножения и имеют лопатчатую форму. То, что похоже на веер на конце брюшка, на самом деле представляет собой множество широких плоских придатков, называемых уроподами. Все эти придатки прикреплены к брюшной стороне Cambarus bartonii , и все они двуветвистые. В целом живот большой и обычно расширенный; однако он может быть согнут под головогрудью. Первый сегмент брюшной полости обычно меньше, чем кзади от него.
Cambarus bartonii имеет открытую систему кровообращения. У него ромбовидное сердце, расположенное кпереди от брюшных сегментов по средней линии спины. Сердце окружено тонким перикардиальным мешком. Прямо перед сердцем лежат гонады. Семенники белые, а яичники оранжевые. Вдоль дорсальной средней линии головогруди лежат сердечный желудок и пилорический желудок. Сразу кзади и сбоку от желудка расположена большая пищеварительная железа.Брюшной нервный тяж лежит под внутренними органами, а мозг находится между стебельками глаз и под ними. (Рассел-Хантер, 1979; Водопич, Мур, 1999)
Репродукция
Спаривание чаще всего происходит весной, но может происходить и летом. Спаривание обычно происходит ночью, потому что шансы встретить друг друга самцом и самкой ночью в девять раз выше.
Размножение предполагает спаривание и может происходить двумя способами.Первый — это отложение спермы в семяприемник у самки. Это происходит, когда сперматозоид самца течет по бороздкам первых плеоподов и попадает в приемник самки. Сперма выходит из самца раков у основания пятой пары ходильных ног через пору. Яйца выпускаются у основания третьей пары ходильных ног. Другая форма размножения включает перенос сперматофора, и в этом случае оплодотворение является внутренним.
В любом случае оплодотворенные яйца сохраняются для созревания на плеоподах самки.Они вылупляются на плеоподах и остаются привязанными к матери вскоре после второй линьки. Говорят, что самка, несущая яйца, находится «в ягоде», потому что масса яиц похожа на ягоду. Самки чаще всего «в ягоде» в мае и июне. (Робертс, 1994; Водопич, Мур, 1999)
Поведение
Cambarus bartonii медленно ходите по дну прудов, ручьев или ручьев в поисках пищи. В случае угрозы они могут стрелять назад, чтобы быстро избежать опасности.Они совершают это быстрое движение назад, сокращая мощные мышцы живота. Cambarus bartonii используют хелипедов для защиты и поиска пищи. Хищники на C. bartonii включают различные виды птиц и черепах. Cambarus bartonii наиболее активен ночью, потому что именно тогда он питается. (Buchsbaum и др., 1987)
Привычки питания
Cambarus bartonii — хищник и падальщик.Он питается разлагающимися органическими остатками, но также ловит мелких животных. Его основными источниками пищи являются улитки, водоросли, личинки насекомых, различные виды червей и головастиков. Он находит пищу на дне источника, в котором обитает, или в почве рядом с водой. (Банистер и Кэмпбелл, 1985)
- амфибии
- яйца
- насекомые
- моллюски
- водные или морские черви
- водные ракообразные
Экономическое значение для людей: положительный результат
Раки — огромная индустрия для многих стран, особенно для Франции и США.Ежегодно потребляется от 400 000 до 800 000 кг (от 880 000 до 1 960 000 фунтов). Ежегодно в искусственных водоемах выращивают 2,6 миллиона фунтов раков. Это создает много рабочих мест для людей, занятых в рыбной отрасли.
Раки полезны и являются источником белка. Сейчас их едят в Соединенных Штатах и во многих странах мира.
Раки также могут иметь лечебные свойства. В Кении — микроскопический кровяной сосальщик.шистом вызывает инфекцию у многих людей. Шистом проникает через кожу и перемещается к мочевому пузырю и другим основным органам, вызывая серьезные повреждения. Ученые выяснили, что личинки шистома вылупляются в пресноводных улитках. Они также обнаружили, что у раков есть аппетит к этим пресноводным улиткам. Размещение большого количества раков (хотя не говорится, какие именно виды будут использоваться; у большинства видов есть аппетит к улиткам) в этих озерах может уменьшить популяцию улиток и, в свою очередь, снизить количество инфекций.
Cambarus bartonii закапывает почву вдоль ручьев, прудов и рек. Это помогает в сельском хозяйстве, потому что перекопка обогащает почву питательными веществами. На многих рисовых фермах раки являются второй культурой, потому что фермеры привозят их после уборки риса, чтобы помочь почве получить много питательных веществ. Затем фермеры собирают раков, когда снова наступает сезон риса. Это не является преимуществом для конкретного вида, потому что в мире существует множество рисовых ферм.
Раки также используются для мониторинга экологического состояния ручьев и рек. В частности, их можно проанализировать на предмет наличия в их тканях определенных загрязнителей. («Раки», 1998; Банистер и Кэмпбелл, 1985)
Экономическое значение для людей: отрицательное значение
Нет никаких известных побочных эффектов Cambarus bartonii на человека.
Статус сохранения
Известно, что этот вид не находится под угрозой исчезновения.
Авторы
Стефани Дженкинсон (автор), Юго-Западный университет, Стефани Фабрициус (редактор), Юго-Западный университет.
Глоссарий
- Неарктика
человек проживает в Неарктической биогеографической провинции, северной части Нового Света. Это включает Гренландию, канадские арктические острова и всю Северную Америку вплоть до юга до высокогорья центральной Мексики.
- двусторонняя симметрия
, имеющий такую симметрию тела, что животное можно разделить в одной плоскости на две зеркальные половины. У животных с двусторонней симметрией есть спинная и вентральная стороны, а также передний и задний концы. Синапоморфия билатериев.
- детрит
частицы органического материала от мертвых и разлагающихся организмов. Детрит — результат деятельности деструкторов (организмов, разлагающих органический материал).
- экзотермический
животных, которые должны использовать тепло, полученное из окружающей среды, и поведенческие адаптации для регулирования температуры тела
- пресная вода
в основном обитает в несоленой воде.
- гетеротермический
имеет температуру тела, которая колеблется в зависимости от температуры окружающей среды; не имеющий механизма или плохо развитый механизм регулирования внутренней температуры тела.
- гибернация
состояние, в которое некоторые животные заходят зимой, когда нормальные физиологические процессы значительно замедляются, что снижает потребность животных в энергии.Акт или условие прохождения зимы в оцепенении или состоянии покоя, обычно предполагающее отказ от гомойотермии у млекопитающих.
- итеропарное
потомков производятся более чем в одной группе (пометы, кладки и т. Д.) И в течение нескольких сезонов (или других периодов, благоприятных для воспроизводства). По определению, итеропородящие животные должны выживать в течение нескольких сезонов (или периодических изменений состояния).
- подвижный
, имеющий возможность перемещаться с одного места на другое.
- натуральный
для плавания
- родной диапазон
район, в котором животное встречается в природе, регион, в котором оно является эндемиком.
- ночной
активен ночью
- всеядное
животное, которое в основном ест всевозможные продукты, включая растения и животных
- яйцекладущие
размножение, при котором яйца выпускает самка; развитие потомства происходит вне организма матери.
- сезонное разведение
разведение привязано к определенному сезону
- малоподвижный
остается на том же участке
- половой
воспроизводство, включающее сочетание генетического вклада двух особей, мужчины и женщины
- наземный
Проживает на земле.
Список литературы
1998. Раки. Всемирная книжная энциклопедия. Лондон: World Book Inc ..
Банистер, К., А. Кэмпбелл. 1985. Омары и пресноводные раки. Стр. 236-239 в Энциклопедии водной жизни. Нью-Йорк: Equinox Ltd.
Buchsbaum, R., M. Buchsbaum, J. Pearse, V. Pearse. 1987. Животные без позвоночника. Чикаго: Издательство Чикагского университета.
Грин, Дж. 1967. Ракообразные. Стр. 172-175 в Энциклопедии жизни животных Ларусс. Нью-Йорк: Книжная компания Mc Graw-Hill.
Макман Л. 1960. Случай строительства дымохода раками * Cambarus bartonii *. Экология, 41: 383-385.
Робертс, Т. 1994. Свет, химические вещества для глаз и некоторые другие факторы как регуляторы общественной активности раков. Экологические монографии, 14: с. 376-378.
Рассел-Хантер, В.1979. Жизнь беспозвоночных. Нью-Йорк: McMillan Publishing Co Inc.
Водопич Д., Мур Р. 1999. Руководство лаборатории биологии. Берр Ридж: У. Х. Фриман и компания.
Раки (от NC WINS) | NCpedia
Камбарус бартони Написано Сарой Пятница
Обновлено Робертом Николсом
Комиссия по ресурсам дикой природы Северной Каролины.
Классификация Класс: Ракообразные
Отряд: Декапода «десятиногая»
Средний размер Около 3 дюймовв длину
Еда Питаются в основном водной растительностью, но также едят червей,
улиток, головастиков и других мелких живых или мертвых существ.
Разведение Большинство дает молодь весной. Самка откладывает сотни яиц; вылупление около 85%. Количество выводков разное. Ученые не уверены, сколько времени требуется для вылупления яиц, но по их оценкам, месяц или больше. Когда яйца вылупляются, появляются крошечные прозрачные версии взрослых раков размером около 1/6 дюйма.долго.
Молодые Молодь цепляется за самку через три линьки, пока не достигнет размера около 1 1/2 дюйма. Раки не проходят личиночную стадию.
Ожидаемая продолжительность жизни Примерно 3-5 лет.
Диапазон и распространение
Некоторые раки являются универсальными и встречаются на большей части территории Северной Каролины, но большинство из них ограничено небольшими географическими районами. Раки-роющие Гринсборо, Cambarus catagius, встречаются только в районе Гринсборо, а колючие раки Северной Каролины, Orconectes carolinensis, встречаются только в бассейнах рек Тар и Нойз на прибрежной равнине.
Общая информация Летом переверните скалу почти в любом ручье в Северной Каролине, и велика вероятность, что рак отскочит от вас назад или даже защитит скалу своими когтями, направленными на вас. Существует по крайней мере 40 различных видов этого обычного пресноводного ракообразного, которые обитают в ручьях, прудах и норах Тар Хил от гор до побережья. Многие из этих видов не встречаются больше нигде на Земле.
Описание Раки выглядят как миниатюрные омары с передней парой сильных щипающих когтей, бронированным телом и широким хвостом.Как и у лобстеров, у раков есть 3 основные части тела: голова, грудная клетка и живот, а также твердый экзоскелет, который защищает их мягкие ткани и органы. Передняя часть тела жесткая, но задняя часть, брюшко или хвост, имеет подвижные сегменты. В области головы 2 пары антенн и крошечные глазки помогают ракам воспринимать окружающую среду. В грудной клетке у раков по 5 ног с каждой стороны (тела) и мягкие перистые жабры, которые используются для дыхания. Первая пара ног — это большие когти или клешни, которые помогают ракам ловить и удерживать добычу.Следующие 4 пары ног — это ходячие ноги; первые 2 пары — это маленькие клешни, используемые для обработки пищи. Под хвостом и после последней пары ходильных ног есть более мелкие и более тонкие придатки, похожие на ноги, называемые пловцами, которые помогают плавать, переносить яйца и удерживать молодых раков. В зависимости от вида окраска раков варьируется от малиново-красного до кобальтово-синего. Некоторые из них могут быть розовыми, коричневыми, зеленовато-черными или даже аквамариновыми.
История и статус Наши биологи ежегодно открывают и описывают новые виды раков, а также документируют распространение экзотических раков в штате.По мере завершения новых исследований биологи продолжают документировать расширение ареала, а также сокращение некоторых раков Северной Каролины.
Среда обитания и привычки
Раки обитают в ручьях, прудах, озерах и болотах по всей Северной Каролине. Обитатели ручьев предпочитают быстро движущиеся и насыщенные кислородом реки и ручьи гор и предгорий. В более медленных ручьях, например на Прибрежной равнине, раки прячутся под камнями и бревнами для защиты.Раков также можно встретить в прудах, озерах и в стоячей воде в придорожных канавах. Некоторые раки зарываются в землю и могут рыть разветвляющиеся туннели глубиной до 15 футов в илистых заболоченных местах. Большинство нор в среднем составляют 2–3 фута в глубину, и их можно найти на приличном расстоянии от стоячей воды. Раки-роющие, обитающие в основном на Прибрежной равнине и в восточной части Северной Каролины, во время копания часто выбрасывают глиняные «дымоходы».
Раки становятся наиболее активными с наступлением темноты или на рассвете и выходят из своих укрытий, чтобы покормиться.Эти стойкие ракообразные питаются в основном водной растительностью, но они также могут есть мелких, живых или мертвых животных, таких как черви, улитки, головастики и рыба. Раки часто становятся добычей других животных, таких как лягушки, черепахи, окуни, еноты, болотные птицы и ястребы. Но раки — бойцы. Лучшая защита — мощный хвост, который отбрасывает раков назад. Если они теряют коготь или ногу в бою, на их месте вырастает новый.
Большинство видов раков в Северной Каролине дают потомство ранней весной, когда много еды и дождя.Некоторые виды размножаются зимой. Для спаривания самец рака хватает самку и переворачивает ее так, чтобы брюшная сторона находилась вместе. Затем самец передает сперму самке через свою первую пару плавников, называемых гоноподом. При оплодотворении сотни крошечных яиц прикрепляются к плавникам самки с липкими выделениями, называемыми яицами. Когда к самке раков прикрепляются оплодотворенные яйца, говорят, что она в ягоде. Более крупные самки обычно откладывают больше яиц.
В отличие от многих других ракообразных, раки не проходят личиночную стадию, а линяют около 6 раз в первые 7-8 месяцев жизни.Линька или выпадение панциря необходимо, потому что раки перерастают свою броню. Они продолжают линять более медленными темпами всю оставшуюся жизнь. Когда они линяют, старый скелет отпадает, и раки с мягким панцирем быстро растут, так как поглощают большое количество воды. Незащищенные, они прячутся после линьки и менее агрессивны, пока через несколько дней не затвердеет новый скелет. Раки прекрасно себя чувствуют в незагрязненной воде, и их количество является хорошим признаком хорошей экологии.
Взаимодействие с людьми Раки ценятся не только с экологической точки зрения, но и с экономической точки зрения.Более крупные раки, такие как красные болотные раки, считаются деликатесом каджунской кухни. Раки также используются в качестве приманки для рыбной ловли, и это привело к некоторым случайным проникновениям неместных раков в новые районы. Неместные раки вредят нашей экосистеме, конкурируя с местными видами тархилов и уничтожая водные растения, которые служат пищей и убежищем для местных животных. ПОЖАЛУЙСТА, НЕ выпускайте раков в водные пути Северной Каролины !!! Попав сюда, их невозможно искоренить! Вот совет для тех, кому не посчастливилось попасть не на тот конец раков.У них есть острый укус, но он укусил их только в качестве последней попытки самообороны. Если им дадут шанс сбежать, они это сделают. Так что, если вы когда-нибудь обнаружите, что к вам прицепились раки, просто
воткните пораженную часть анатомии обратно в воду, где раки будут чувствовать себя в большей безопасности и уплывут. Этот метод также работает с черепахами.
Примечание: «раки» относятся к одному виду или отдельному экземпляру, тогда как «раки» относятся ко многим видам.
Судьба золотистых желез и нерестовое поведение у неместных раков: сравнение гонохорических речных раков и партеногенетических мраморных раков.
Abstract
В период до нереста у самок пресноводных раков развиваются золотистые железы на нижней стороне плеона.Эти железы производят слизь для временной структуры, похожей на палатку, в которой яйца оплодотворяются и прикрепляются к плеоподам. Долгосрочное наблюдение за самками рака Procambarus fallax , размножающихся двуполым путем, содержащихся в неволе, показало, что голые железы развиваются в конце зимы и в конце лета каждого года независимо от присутствия самцов. Однако, в отличие от спарившихся самок, самки без спаривания не образуют ни оплодотворяющей палатки, ни нереста. Их голые железы сохранялись необычно долго и исчезли только во время следующей линьки.Судя по всему, женщины используют информацию о наличии сперматозоидов либо для воспроизводства, либо для сохранения ресурсов. Мраморный рак, Procambarus virginalis , партеногенетический потомок слепого рака, развил голые железы примерно в одни и те же периоды года, но нерестился, несмотря на отсутствие спаривания. Эти находки указывают на то, что на пути от гонохоризма к партеногенезу регуляция активности голых желез и нереста была отделена от спаривания. Таким образом, пара видов Procambarus fallax / Procambarus virginalis кажется особенно подходящей для исследования физиологических, молекулярных и генетических механизмов, лежащих в основе нереста пресноводных раков.
Введение
За несколько недель до нереста у пресноводных раков на нижней стороне плеона развиваются выступающие белесые блестящие железы (Reynolds 2002). Эти железы сначала выглядят как слабые кремово-беловатые пятна и становятся все более и более отчетливыми, что является хорошим индикатором предстоящего нереста (Jussila et al. 1996). Незадолго до откладывания яиц голые железы выделяют полупрозрачную слизь, которая образует тентовую структуру на нижней стороне грудной клетки и плеона (Andrews 1904; Vogt 2016).В этой студенистой массе сперматозоиды мобилизуются из сперматофоров, которые прикреплены или хранятся внутри, в зависимости от семейства раков (Reynolds 2002; Vogt 2002; Niksirat et al. 2013). Палатка для оплодотворения дополнительно обеспечивает прикрепление оплодотворенных яиц к плеоподам, где они высиживаются в течение нескольких недель до вылупления первой молоди и далее (Vogt and Tolley 2004). Глэровые железы и палатка оплодотворения являются уникальными особенностями пресноводных раков и встречаются во всех трех семействах: Astacidae, Cambaridae и Parastacidae (Andrews 1906; Mason 1970; Thomas and Crawley 1975; Jussila et al.1996; Фогт 2016). Более простые цементные железы, которые только способствуют прикреплению яиц к плеоподам, встречаются и у других Decapoda (Adiyodi and Anilkumar 1988).
Во время конференций по ракообразным и студенческих курсов я часто сталкивался с вопросом, что происходит с желчными железами и зрелыми ооцитами, если самки раков не спариваются. К сожалению, нет опубликованных данных о судьбе желчных желез и очень мало данных о нересте у несвязанных самок (Woodlock, Reynolds 1988; Reynolds et al.1992; Холдич и др. 1995; Tropea et al. 2010; Buřič et al. 2011). Одна из возможностей состоит в том, что несвязанные самки нерестятся как спарившиеся самки, но неоплодотворенные яйца разлагаются. В качестве альтернативы слизь и зрелые ооциты могут реабсорбироваться, чтобы избежать растраты ресурсов. В настоящем сообщении я сообщаю о судьбе головых желез и нерестовом поведении у выращенных в лаборатории спарившихся и не спарившихся речных раков Procambarus fallax и их партеногенетически воспроизводящегося родственника, мраморного рака Procambarus virginalis .У мраморных раков нет самцов, и они произошли от речных раков в результате аутотриплоидизации и сопутствующего перехода от гонохоризма к партеногенезу (Scholtz et al. 2003; Martin et al. 2010; 2016; Vogt et al. 2015). Недавно они были признаны отдельным видом из-за репродуктивной и географической изоляции от Procambarus fallax , уникальных генетических и эпигенетических признаков, а также превосходного роста и плодовитости (Vogt et al. 2015).
Материалы и методы
Самки речного рака Procambarus fallax (Hagen 1870) (Decapoda: Cambaridae) были отделены от своих сородичей-самцов на пятой стадии молоди, первой стадии жизни, которая позволяет определить пол по внешнему признаку.В последующие пять месяцев их держали вместе в женской группе, а затем по отдельности. Три из этих самок остались без спаривания, а еще три самки были повязаны с самцами в течение сезонов размножения весной 2015, осенью 2015 и весной 2016. Спаривание производилось путем спаривания отдельных самок с гладкими железами и отдельных самцов с крючками на седалищных костях 3-й и 3-й седалищных желез. 4-е переоподы (показатели половой зрелости) в контейнере 30 × 25 × 20 см. Копуляция считалась успешной, когда самцы переворачивали самок на спину и вставляли их копулятивные органы в приемник спермы (annulus ventralis) самки не менее чем на 30 минут.Все спарившиеся и не спарившиеся самки содержались индивидуально в пластиковых контейнерах 30 × 25 × 20 см, снабженных гравием и укрытием. В качестве источника воды использовалась водопроводная вода, которую меняли раз в неделю. Температура воды колебалась от ~ 15 ° C зимой до ~ 25 ° C летом, и фотопериод был естественным режимом окружающей среды Гейдельберга (Германия). Раков кормили ежедневно ad libitum гранулами TetraWafer Mix. Период наблюдений длился с 1 июля 2014 г. по 31 мая 2016 г.Я дал время в неделях только потому, что в начале созревания слепой железы трудно решить, началось ли уже развитие или нет. Данные по Procambarus fallax были сопоставлены с моими записями о наличии блестящих желез у партеногенетических мраморных раков Procambarus virginalis , которые я собирал за последние 13 лет. Мраморных раков выращивали в тех же условиях, что и речных раков, индивидуально или коллективно.
Результаты
У самок обоих видов раков в период размножения развиваются голые железы. Эти железы были расположены в последней грудной части грудины, грудины и плевре плеона, уропод и плеопод (рис. 1А, В). Головые железы развивались автономно, в отсутствие самцов.
Рис. 1. Глеровские железы и приемник для сперматозоидов у раков. (A) Зрелые железы голени на нижней стороне плеона партеногенетической самки Procambarus virginalis .Беловатые железы расположены в грудины (стрелка), плевре (наконечник стрелки), уроподах (u) и плеоподах (p). Пруток: 5 мм. (B) Плеон самки Procambarus fallax без спаривания с обнаженными железами через 12 недель после их первого появления. Пруток: 2 мм. (C) Сканирующая электронная микрофотография брюшного кольца самки Procambarus virginalis . Бар: 200 мкм.
В трех проанализированных периодах воспроизводства нерестовые самки Procambarus fallax не спаривались, хотя у них регулярно развивались голые железы (Таблица 1).Напротив, спарившиеся самки обычно откладывали яйца. В последнем случае блестящие железы были видны в течение трех-пяти недель. Их опорожняли незадолго до кладки яиц, чтобы сформировать палатку для оплодотворения. У несвязанных самок золотистые железы сохранялись до 12 недель (таблица 1) и исчезли только во время следующего шелушения.
Таблица 1. Судьба золотистых желез и нерестовое поведение у не спарившихся и не спарившихся Procambarus fallax самок.
Мраморных раков, выращиваемых в лаборатории, в основном воспроизводятся два раза в год, как и речных раков (Vogt 2015).Некоторые особи размножаются только один-три раза в год. За 3-6 недель до нереста у них были обнаружены железы с увеличивающейся интенсивностью окрашивания. В отличие от несвязанных самок речного рака, партеногенетические самки мраморных раков производили палатки для оплодотворения и нерестились. Только в одном из 103 задокументированных случаев самка с голыми железами пропустила нерест и вместо этого выполнила линьку. Золотистые железы этого экземпляра исчезли во время линьки, как и у несвязанных раков.
Обсуждение
Сравнение не спарившихся самок гладкоплодных раков и партеногенетических мраморных раков выявило интересные различия в отношении нереста и судьбы гладких желез.Спаривающиеся самки и самки мраморных раков обычно нерестятся дважды в год (Vogt 2015). Эти нереста связаны с опорожнением желёз. У слабых раков оба события зависят от спаривания. Несвязанные Procambarus fallax самок не порождают и не опорожняют свои голые железы. Вместо этого блестящие железы сохраняются еще много недель, вероятно, в ожидании более позднего шанса спаривания. Это контрастирует с мраморными раками, у которых развиваются голые железы и нерестятся в отсутствие самцов.Очевидно, что информационная система, позволяющая различать условия спаривания и отсутствия спаривания, была потеряна или изменена во время перехода от гонохоризма к партеногенезу в паре видов Procambarus fallax / Procambarus virginalis .
Полное подавление нереста наблюдалось также у самок астцид Austropotamobius pallipes , Astacus leptodactylus и Pacifastacus leniusculus по данным Holdich et al.(1995). Напротив, Вудлок и Рейнольдс (1988) наблюдали нормальный нерест, но распад прикрепленных яиц у несвязанных самок Austropotamobius pallipes , которые содержались в парах самок. У самок выращенных в лабораторных условиях Cherax destructor (Parastacidae), не имеющих спаривания, нерестовое поведение было весьма разнообразным: около 20% нерестились при выращивании при 30 ° C, но не появлялись при 27 ° C (Tropea et al. 2010). Нерест не спарившихся самок произошел и у факультативно партеногенетических самок камбарид Orconectes limosus (Buřič et al.2011, 2013). Из 30 не спаривавшихся партеногенетических самок 28 нерестились, и только одна самка не отнерестилась (оставшаяся особь погибла) (Buřič et al. 2011), как и у мраморных раков. Причины различий в нерестовом поведении самок, не имеющих спаривания, между указанными размножающимися половым путем видами и внутри них неизвестны, но могут зависеть от видовых различий, условий окружающей среды, предэкспериментального опыта и статуса питания особей. Информация о судьбе желёзок отсутствует во всех цитированных выше сообщениях.Подавление секреции и нереста гололеда можно интерпретировать как энергосберегающую стратегию.
Созревание ооцитов не зависит от присутствия самцов, как показали все вышеупомянутые исследования. То же самое касается созревания желёзок, которые развиваются за несколько недель до нереста. Оба процесса, по-видимому, тесно связаны и вызываются факторами окружающей среды, в основном температурой и световым периодом (Reynolds 2002; Vogt 2015), которые влияют на гормональную систему. Созревание яичников ингибируется гормоном, ингибирующим вителлогенез (VIH) из системы X-орган-синусовая железа, и стимулируется гормоном, стимулирующим яичники (OSH), из ганглиев грудных нервов (Reynolds 2002; Vogt 2002).Созревание блестящих желез, вероятно, регулируется теми же гормонами.
Спаривание может быть одним из спусковых механизмов для нереста, как предполагает Рейнольдс (2002). Это предположение подтверждается экспериментами Holdich et al. (1995), которые спарили самок с самцами другого вида. Такие самки обычно нерестились, но икринки не развивались. Однако в естественных условиях между спариванием и нерестом часто бывает много дней или даже недель, что позволяет предположить, что нерест также может зависеть от воспоминаний о спаривании самкой или ощущения присутствия спермы.У астацидных и парастакидных раков сперматофоры прикреплены снаружи к грудным пластинам грудины и хвостовому вееру (Reynolds 2002; Vogt 2002; Niksirat et al.2013), и, таким образом, самки могут легко ощущать присутствие сперматозоидов своими переоподами. У Cambaridae сперма хранится внутри вентрального кольца (рис. 1C) (Reynolds 2002; Vogt et al. 2004), и в этом случае механорецепторы могут дать информацию о состоянии ее наполнения. №
Неизвестно, могут ли раки добровольно опорожнять железы и яичники за счет сокращения мускулатуры желез и яичников.Более вероятно, что оба процесса являются непроизвольными и регулируются гормонами, такими как простагландин F 2α , который, как было показано, резко усиливается на заключительной стадии оогенеза у Procambarus paeninsulanus (Spaziani et al. 1995). Тесты in vitro показали, что этот гормон вызывает сокращение ткани яичников в зависимости от дозы.
Настоящее исследование является первым отчетом о судьбе железистой оболочки без спаривания раков, выращенных в контролируемых условиях.
Парные противники были помещены в | $ 30,5 {\ text {cm width}} \ times 40.6 {\ text {cm length}} \ times 14.6 {\ text {cm depth}} $ | пластиковые емкости, наполненные свежей фильтрованной водопроводной водой; внутри танков противники были разделены прозрачными перегородками и получили возможность акклиматизироваться за | $ 2 \, {\ rm {min}} $ | . Затем разделители были сняты, и агонистические взаимодействия были записаны на видео для | $ 10 \, {\ rm {min}} $ | . По завершении каждого испытания обоих животных возвращали в их резервуары для хранения.Все резервуары тщательно промывались и наполнялись свежей фильтрованной водой между испытаниями. Видеозаписи испытаний оценивались слепым рецензентом за время, проведенное в физических агонистических взаимодействиях, обозначенных как «схватка» и определяемых как бокс, удары, хватание или толкание чела одним или обоими раками; и для проявления поведения, указывающего на различные уровни агрессивного взаимодействия (Таблица 1). Общее время, потраченное на «захватное» поведение, и доля схваток, достигших максимальной интенсивности, сравнивали между обработками как меры общего агонистического вложения.
Таблица 1Этограмма окончательного поведения, используемая для приблизительного определения уровней интенсивности боя. Все виды поведения оценивались для обоих раков в паре. Этограмма адаптирована из Bruski & Dunham (1987) для использования в текущем исследовании.
| Уровень интенсивности . | Описание . |
|---|---|
| 0 | Нет взаимодействия |
| 1 | Приближение — приближение на одну длину тела противника, приводящее к отступлению противника в любом направлении |
| 2 | Выпад / угроза — быстрое приближение противника с Чела, поднятые на дисплее угрозы |
| 3 | Захват — бокс, удар, хватание или толкание чела одним или обоими раками |
| 4 | Переворот — раки повернуты на бок или спиной, когда их схватил с чела соперника |
| Уровень интенсивности . | Описание . |
|---|---|
| 0 | Нет взаимодействия |
| 1 | Приближение — приближение на одну длину тела противника, приводящее к отступлению противника в любом направлении |
| 2 | Выпад / угроза — быстрое приближение противника с Челы, поднятые на дисплее угрозы |
| 3 | Захват — бокс, удар, хватание или толкание чела одним или обоими раками |
| 4 | Переворот — раки повернуты на бок или спиной, когда их схватил с чела оппонента |
Этограмма окончательного поведения, используемая для приблизительного определения уровней интенсивности боя.Все виды поведения оценивались для обоих раков в паре. Этограмма адаптирована из Bruski & Dunham (1987) для использования в текущем исследовании.
| Уровень интенсивности . | Описание . |
|---|---|
| 0 | Нет взаимодействия |
| 1 | Приближение — приближение на одну длину тела противника, приводящее к отступлению противника в любом направлении |
| 2 | Выпад / угроза — быстрое приближение противника с Чела, поднятые на дисплее угрозы |
| 3 | Захват — бокс, удар, хватание или толкание чела одним или обоими раками |
| 4 | Переворот — раки повернуты на бок или спиной, когда их схватил с чела соперника |
| Уровень интенсивности . | Описание . |
|---|---|
| 0 | Без взаимодействия |
| 1 | Приближение — приближение на одну длину тела к противнику, приводящее к отступлению противника в любом направлении |
| 2 | Выпад / угроза — быстрое приближение противника с Челы, поднятые на дисплее угрозы |
| 3 | Захват — бокс, удар, хватание или толкание чела одним или обоими раками |
| 4 | Переворот — раки повернуты на бок или спиной, когда их схватил с чела оппонента |
11″ data-legacy-id=»s3″> Результаты Не было значительной разницы в общем размере самок между падениями | $ (n = 34, \, \ bar {x} = 35.203 {\ text {mm}}) $ | и пружина | $ (n = 28, \, \ bar {x} = 34.993 {\ text {mm}}, \, t = 0.219, \, {\ text {DF}} = 60, \, P = 0.827 ) $ | , либо для женщин в маленьком | $ ({\ text {fall:}} = 11, \, \ bar {x} = 30.572 {\ text {mm}} $ | ; | $ {\ текст {весна:}} n = 11, \, \ bar {x} = 31,046 {\ text {mm}}, \, t = -0,555, \, {\ text {DF}} = 20, \, P = 0,585 $ | ) или большой | $ ({\ text {fall:}} n = 23, \, \ bar {x} = 37,417 {\ text {mm}}; \, {\ text {spring:}} n = 17, \, \ bar {x} = 37,547 {\ text {mm}}, \, t = — 0,214, \, {\ text {DF}} \, = 38, \, P = 0.832) $ | размерные классы. Наблюдалась значительная разница в общем размере мужчин между падениями | $ (n = 34, \, \ bar {x} = 35.247 {\ text {mm}}) $ | и пружина | $ (n = 42, \, \ bar {x} = 33,136 {\ text {mm}}, \, t = 2,867, \, {\ text {DF}} = 74, \, P = 0,005 ) $ | , но не для маленьких | $ ({\ rm {fall}}: \, \, n = {\ rm {14}}, \, \, \ bar x = {\ rm {32} } .000 \, \, {\ rm {mm}}; \, \, {\ rm {spring}}: \, \, n = {\ rm {24}}, \, \, \ bar x = { \ rm {3}} 0. {\ rm {746}} \, \, {\ rm {mm}}, \, \, t = {\ rm {\ bar 2}}. {\ rm {376}} , \, \, {\ rm {DF}} = {\ rm {36}}, \, \, P = 0.0 {\ rm {23}}) $ | или большой | $ ({\ text {fall:}} n = 20, \, \ bar {x} = 37,520 {\ text {mm}}; \, {\ text {spring:}} n = 18, \ , \ bar {x} = 36,322 {\ text {mm}}, \, t = 2,275, \, {\ text {DF}} = 36, \, P = 0,029) $ | группы отдельно.
При сравнении времени, затраченного на общее агонистическое поведение, и лечение, и размер показали значительный эффект в различных сравнениях (таблица 2, рис. 1). Самцы | $ (n = 17) $ | и женщины | $ (n = 22) $ | в осеннем репродуктивном сезоне было обнаружено значительное влияние секса на общее время, затрачиваемое на агонистическое поведение, при этом самцы тратят больше времени на такое поведение, чем самки, и отсутствие значительного влияния размера на общее агонистическое поведение в этих двух группах.Существенной разницы между мужчинами | $ (n = 21) $ | и женщины | $ (n = 19) $ | летом, в период отсутствия репродукции, общее время, проведенное в агонистическом поведении, но и самцы, и самки для этого сравнения показали значительный эффект размера, при этом более крупные животные тратят значительно больше времени на агонистическое поведение, чем более мелкие животные. Сравнение репродуктивных и не репродуктивных особей не показало значительных различий в общем времени, проведенном в агонистическом поведении в зависимости от сезона или размера для любого пола, хотя для женщин взаимодействие между репродуктивным состоянием и размером имело значительный эффект и сохранялось в модель (но которая не имела значения при исправлении Бонферрони | $ {\ rm {\ alpha}} = 0.0125 $ | ).
Таблица 2 Результаты дисперсионного анализа с фиксированными эффектами сезона, например, репродуктивные самцы по сравнению с непродуктивными самцами, или пол, например, репродуктивные самцы по сравнению с репродуктивными самками. Во всех сравнениях размер был включен как фактор; член взаимодействия был незначительным | $ (P> 0,05) $ | во всех случаях, кроме сравнения репродуктивных самок с непродуктивными самками, и был исключен из анализов, в которых он не был значимым.
. Фактор 1 (размер)
. Фактор 2 (сезон или пол)
. Фактор 1 * фактор2
. Контрастность
. ф
. | $ P $ |
. ф
. | $ P $ |
. ф
. | $ P $ |
. Между сезонами: 1.Репродуктивные самцы и непродуктивные самцы 0,468 0,498 1,174 0,286 — — 2. Репродуктивные самки и непродуктивные самки 4,820 0,034 0,146 4,027 0,052 Между полами: 3.Репродуктивные самцы и репродуктивные самки 0,065 0,801 9,071 0,005 * — — 4. Непродуктивные самцы и непродуктивные самки 7,381 9 * 0,025 0,876 — —
. Фактор 1 (размер)
. Фактор 2 (сезон или пол)
. Фактор 1 * фактор2
. Контрастность
. ф
. | $ P $ |
. ф
. | $ P $ |
. ф
. | $ P $ |
. Между сезонами: 1.Репродуктивные самцы и непродуктивные самцы 0,468 0,498 1,174 0,286 — — 2. Репродуктивные самки и непродуктивные самки 4,820 0,034 0,146 4,027 0,052 Между полами: 3.Репродуктивные самцы и репродуктивные самки 0,065 0,801 9,071 0,005 * — — 4. Непродуктивные самцы и непродуктивные самки 7,381 9 * 0,025 0,876 — —
Таблица 2 Результаты ANOVA с фиксированными эффектами сезона, например, репродуктивные самцы по сравнению с непродуктивными самцами или полом, e.g, репродуктивные самцы по сравнению с репродуктивными самками. Во всех сравнениях размер был включен как фактор; член взаимодействия был незначительным | $ (P> 0,05) $ | во всех случаях, кроме сравнения репродуктивных самок с непродуктивными самками, и был исключен из анализов, в которых он не был значимым.
. Фактор 1 (размер)
. Фактор 2 (сезон или пол)
. Фактор 1 * фактор2
. Контрастность
. ф
. | $ P $ |
. ф
. | $ P $ |
. ф
. | $ P $ |
. Между сезонами: 1. Репродуктивные самцы по сравнению с непродуктивными самцами 0.468 0,498 1,174 0,286 — — 2. Репродуктивные самки по сравнению с непродуктивными самками 4,820 0,034 2,207 0,146 4,0 Между полами: 3. Репродуктивные самцы и репродуктивные самки 0.065 0,801 9,071 0,005 * — — 4. Непродуктивные самцы и непродуктивные самки 7,381 0,010 * 76 — —
. Фактор 1 (размер)
. Фактор 2 (сезон или пол)
. Фактор 1 * фактор2
. Контрастность
. ф
. | $ P $ |
. ф
. | $ P $ |
. ф
. | $ P $ |
. Между сезонами: 1.Репродуктивные самцы и непродуктивные самцы 0,468 0,498 1,174 0,286 — — 2. Репродуктивные самки и непродуктивные самки 4,820 0,034 0,146 4,027 0,052 Между полами: 3.Репродуктивные самцы и репродуктивные самки 0,065 0,801 9,071 0,005 * — — 4. Непродуктивные самцы и непродуктивные самки 7,381 9 * 0,025 0,876 — —
Мы обнаружили аналогичные эффекты размера и лечения, когда сравнивали долю драк, достигших максимального уровня интенсивности.Во всех случаях, кроме одного (самки летом, нерепродуктивный сезон, | $ P = 0,018 $ | ), не было существенной разницы в доле испытаний, достигающих максимальной интенсивности между группами малого и большого размера (репродуктивные самцы | $ P = 0,153 $ | , репродуктивные самки | $ P = 0,364 $ | , непродуктивные самцы | $ P = 0,367 $ | ). Следовательно, для трех последних курсов лечения были объединены большие и малые группы. Как и в случае продолжительности агонистических взаимодействий, значительно больше драк между репродуктивными самцами достигли максимальной интенсивности, чем между репродуктивными самками | $ (P = 0.001) $ | . Не было обнаружено существенной разницы в доле, достигающей максимальной интенсивности между непродуктивными самцами и крупными не репродуктивными самками летом | $ (P = 0,721) $ | ; тем не менее, значительно больше драк между не репродуктивными самцами достигло максимальной интенсивности, чем между маленькими не репродуктивными самками | $ (P = 0,003) $ | . Точно так же значительно больше испытаний между крупными непродуктивными самками достигло максимальной интенсивности, чем испытания между репродуктивными самками | $ (P = 0,020) $ |, в то время как не было значительной разницы в интенсивности между маленькими непродуктивными самками и репродуктивными самками | $ ( Р = 1.000) $ | . Не было существенной разницы в доле испытаний, достигающих максимальной интенсивности между самцами в репродуктивном и не репродуктивном периодах | $ (P = 0,743) $ | .
Обсуждение
Существует множество факторов, как внутренних (размер тела и чела, пол, репродуктивное состояние, предыдущий социальный опыт, мотивационное состояние, нейрохимия), так и внешних (факторы общения с окружающей средой, воспринимаемая ценность ресурсов), которые влияют на количество времени и риски для индивидов. готовы вкладывать средства в агонистические взаимодействия (Мур, 2007).Наши данные показывают, что самцы O. quinebaugensis проводили значительно больше времени в агонистическом поведении и участвовали в большей части драк с максимальной интенсивностью, чем самки в осеннем репродуктивном сезоне, как мы и предсказывали. Это может указывать на то, что самцы реагируют на половой отбор, вкладывая больше энергии в агонизм, если доминирующие самцы имеют репродуктивное преимущество.
Если доминирование действительно связано с повышенной приспособленностью самцов, можно ожидать, что в течение репродуктивного сезона, когда этот ресурс является наиболее ценным, будет инвестировано больше времени и, следовательно, больший риск.Другие исследования показали, что интенсивность и продолжительность агонистических встреч коррелируют с ценностью ресурса и его доступностью для участников (Bergman and Moore, 2003; Stocker and Huber, 2001; обзор в Moore, 2007). У раков, например, материнские самки (несущие икру или молодые особи) могут быть намного более агрессивными, чем нематериальные самки (Figler et al., 1995b), весной, когда качество и выживаемость молоди напрямую влияют на приспособленность самок. В нашем эксперименте, однако, не было значительной разницы во времени, затраченном на агонизм, или пропорции схваток, достигающих максимальной интенсивности, между репродуктивными и не репродуктивными самцами.В частности, в классах большого размера для анализа продолжительности самцы Формы II проводили столько же времени в агрессии, как и самцы Формы I. Точно так же крупные непродуктивные самки проводили больше времени в агрессии и имели более высокую долю драк с максимальной интенсивностью, чем репродуктивные самки. Хотя эта разница была значимой в отношении интенсивности борьбы, она не была значимой для времени, проведенного в агонистическом поведении, вероятно, в результате сильного эффекта размера у не репродуктивных самок.В целом, в то время как поведение репродуктивных самцов и самок согласуется с нашей гипотезой о том, что половой отбор стимулирует инвестиции в агонизм, данные по нерепродуктивным ракам — нет. Мы предлагаем два альтернативных объяснения этого наблюдения.
Во-первых, летний сезон представляет собой основной период роста как молоди, так и взрослых раков O. quinebaugensis ; такие сезонные закономерности линьки и роста обычны для раков умеренного климата (Reynolds, 2002).В этих условиях ценность таких ресурсов, как еда и жилье, может возрасти как для мужчин, так и для женщин. Монополизация пищевых ресурсов может привести к ускоренному росту и улучшению индивидуальной физической формы (Fero et al., 2007), а размер часто положительно коррелирует с результатами борьбы раков (например, Pavey and Fielder, 1996; Daws et al., 2002). ; Schroeder, Huber, 2001; Figler et al., 1995a; Klocker and Strayer, 2004; обзор в Moore, 2007) и плодовитость у самок (Rubolini et al., 2006; Reynolds, 2002), что потенциально может привести к репродуктивным преимуществам на более позднем этапе жизненного цикла для обоих полов. Укрытия также могут быть ресурсом равной ценности, который потребовал бы увеличения агонистических вложений для обоих полов, поскольку частая линька также увеличивает долю времени, в течение которого рак становится более уязвимым для хищников (Hamr, 2002). Следовательно, летом мужчины и женщины могут одинаково вкладывать средства в агонизм, потому что наиболее ценные ресурсы одинаково важны для обоих полов. Осенью относительная важность ресурсов может измениться в ответ на замедление роста и охлаждение температуры воды, а инвестиции в конкуренцию за партнеров, а не за еду или жилье отражаются в ориентированных на пол вложения в агонизм.
В качестве альтернативы, самки во время репродуктивного сезона могут извлечь большую пользу из воздержания от этих увеличенных агонистических вложений в пользу других энергетических потребностей. Для мужчин было высказано предположение, что инвестиции в агонистические взаимодействия должны конкурировать с инвестициями в другие компоненты, связанные с приспособленностью, такие как привлечение партнера или родительская забота (Qvarnström, 1997; Griffith and Sheldon, 2001), и можно было бы ожидать аналогичной торговли. снижение расхода энергии для женщин (например,г., Ветцель, 2002; Бернардо, 1996). Первичная репродуктивная деятельность самок в это время заключается в формировании яиц (Galeotti et al., 2006), что, вероятно, требует больших затрат энергии. Самки могут получить больше преимуществ в фитнесе, вкладывая больше средств в производство яиц, а не борясь за доступ к партнерам или пище, поскольку репродуктивный успех самок ограничен количеством яиц, которые она производит, а также качеством и выживаемостью ее потомства (Trivers, 1972). Было высказано предположение, что размер яйца является решающим фактором, определяющим приспособленность потомства (Bernardo, 1996), а вложение энергии в схватку с аналогами может ограничить способность самки производить большие яйца или кладку.
Поиск партнера и дискриминация также могут повлечь за собой энергетические затраты для женщин (Kokko et al., 2002). Инвестиции в агонизм в репродуктивном сезоне также могут отвлекать от времени и энергии, которые самки могут тратить на отбор и спаривание желаемых, высококачественных самцов, что в конечном итоге может повлиять на качество их потомства (Kokko et al., 2002; Berglund et al. ., 1996). У таких видов, как O. quinebaugensis , у которых самки хранят сперму и откладывают оплодотворение до нескольких месяцев после спаривания (Hamr, 2002), самки также могут получить косвенные выгоды от выделения времени и энергии на получение нескольких спариваний (Jennions and Petrie, 2000).Любые травмы, полученные в результате агрессивного поведения во время репродуктивного сезона, также могут помешать самкам спариваться с желаемым самцом или вообще успешно спариваться.
Весной, когда самка вкладывает средства в потомство в виде эмбрионов и частично независимых молодых особей, самки могут извлечь выгоду за счет повышенной агрессии, если это позволяет им оставаться в приюте, предположительно как компонент защиты потомства (Фиглер). и другие., 2001). Мур (2007) в своем обзоре соответствующей литературы обнаружил, что самки матери были более агрессивными, чем самки нематериального происхождения и репродуктивные самцы, а Фиглер и др. (2001) предположили, что это было частично связано с внутренним состоянием, поскольку даже самки матери с удаленным потомством показали более высокую защиту укрытия по сравнению с самками нематериального происхождения при сравнении предыдущих исследований. В совокупности это говорит о том, что вклад женщин в агонистические взаимодействия может происходить в рамках некоторой формы полового отбора, но в другом направлении, чем можно было бы ожидать от мужчин.
Мы также сообщаем противоречивые данные о влиянии размера на агонистическое поведение. Летом наблюдалось значительное влияние размерной группы как для самцов, так и для самок в зависимости от продолжительности: более крупные раки обоих полов проводили больше времени в агонистическом поведении, чем более мелкие раки. Также наблюдалось влияние размера на интенсивность агрессии между самками летом: у крупных самок была более высокая доля драк, достигающих максимальной интенсивности, чем у маленьких самок. Принято считать, что размер является основным фактором, который обычно предсказывает исход и агрессивное состояние при взаимодействии раков (Pavey and Fielder, 1996; Daws et al., 2002; Клокер и Страйер, 2004; Figler et al., 1995a; Шредер и Хубер, 2001; рассмотрено в Moore, 2007), и было высказано предположение, что маленькие и большие раки могут применять разные стратегии борьбы в зависимости от ценности ресурсов и риска для человека (Schroeder and Huber, 2001). Более крупные, более доминирующие люди потенциально меньше рискуют вступить в агонистическое поведение, поскольку более вероятно, что они добьются успеха, особенно в паре с меньшим противником. Однако мы не наблюдали подобных эффектов размера в брачный период, когда и большие, и маленькие особи обоих полов одинаково вкладывали средства в драки.
Хотя объяснение этому наблюдению неясно, мы отмечаем, что в нерепродуктивной коллекции животных некоторые (возможно, многие) особи обоих полов в группе меньшего размера, возможно, никогда не претерпели периода воспроизводства, в то время как особи в группах большего размера вероятность воспроизводства выше в предыдущем году. Этим можно объяснить значительную разницу, наблюдаемую в доле схваток, достигающих максимальной интенсивности, между репродуктивными самками осенью и крупными непродуктивными самками летом, в то время как сравнение репродуктивных самок осенью и мелких непродуктивных самок летом было незначительным.Материнские самки других видов раков проявляют повышенную агрессию по отношению как к репродуктивным самцам, так и к нематеринским самкам (Figler et al, 1995b), и предполагается, что это частично связано с изменением внутреннего состояния (Figler et al. ., 2001). Следовательно, возможно, что у крупных самок летом мог быть некоторый остаточный внутренний эффект, связанный с недавно рассеяной кладкой молоди, тогда как у маленьких самок меньше шансов иметь недавнюю кладку, хотя недавнее материнское состояние большинства самок в летняя группа была неизвестна.
В целом, наши данные показывают, что индивидуальные вложения в агонизм особей O. quinebaugensis различаются в зависимости от сезона, пола и размера. Тем не менее, мы специально выдвинули гипотезу о том, что взаимодействия с доминированием являются сексуально отобранным поведением, и предсказали, что репродуктивные самцы будут проявлять повышенную агрессию по сравнению как с репродуктивными самками, так и с непродуктивными раками, и наши данные не обеспечивают убедительной поддержки этой гипотезы. Вместо этого наши данные показывают, что агонистические взаимодействия у O.quinebaugensis может регулироваться сложным и динамичным набором индивидуальных затрат и выгод, и это дополнительное экспериментирование может помочь выяснить роль окружающей среды, жизненного цикла и других факторов в социальном поведении этого таксона.
Благодарности
Мы хотели бы поблагодарить Э. Райдер за ее помощь в анализе данных и следующих студентов за их помощь в сборе данных: Л. Амендола, Э. Гринуэй, К. Рикер, Р.Стэнфорд, У. Терротт, Т. Малхерн и М. Беккер. Наконец, мы хотели бы поблагодарить Инженерный корпус армии США за предоставление разрешения на специальное использование (Л. М.) для сбора раков на плотине Ист-Бримфилд в Стербридже, Массачусетс.
Список литературы
Berglund
A.
, Bisazza
A.
и Pilastro
A.
1996
Вооружения и украшения: эволюционное объяснение свойств двойной полезности.
Биологический журнал Линнейского общества
58
385
— 399
Бергман
DA
и Мур
PA
2003
Полевые наблюдения за внутривидовым агонистическим поведением двух видов раков Orconectes rusticus и Orconectes virilis , в разных местообитаниях.
Биологический бюллетень
205
26
— 35
Бергман
D.A.
и Moore
P. A.
2005
Продолжительное воздействие социальных запахов изменяет последующие социальные взаимодействия у раков Orconectes rusticus .
Поведение животных
70
311
— 318
Бергман
DA
, Козловски
К.
, Макинтайр
JC
, Хубер
, Р.
, А.G.
и Moore
P. A.
2003
Временная динамика и связь эффектов победителя у раков, Orconectes rusticus .
Поведение
140
805
— 825
Бернардо
J.
1996
Особый материнский эффект размера отростка, особенно размера яйца: модели, модели, качество доказательств и интерпретаций.
Американский зоолог
36
216
— 236
Порода
M.D.
, Smith
S. K.
и Gall
B. G.
1980
Поведенческие стратегии во время агонистических взаимодействий между самцами у тараканов.
Поведение животных
28
1063
— 1069
Кандолин
U.
1999
Соревнования самцов и самцов облегчают выбор самок колюшки.
Труды Лондонского королевского общества B: Биологические науки
266
785
— 789
Кокс
C.R.
и Le Boeuf
B. J.
1977
Подстрекательство самок к мужской конкуренции: механизм полового отбора.
Американский натуралист
111
317
— 335
Daws
AG
, Грили
J.
, Konzen
K.
и Moore
PA
2002
Предыдущий опыт изменяет исход агрессивных взаимодействий между самцами раков, Procambarus clarkii .
Поведение и физиология в морской и пресноводной среде
35
139
— 148
Fero
K.
, Simon
JS
, Jourdie
V.
и Moore
Moore
2007
Последствия социального доминирования для использования ресурсов раков.
Поведение
144
61
— 82
Фиглер
M.H.
, Finkelstein
JE
, Twum
M.
и Peeke
HVS
1995a
Самцы красных болотных раков, Procambarus clarkii , сразу преобладают более мелкие представители традиционных сообществ. , смешанный сородич.
Агрессивное поведение
21
225
— 236
Фиглер
M. H.
, Twum
M.
, Finkelstein
JE
и Peeke
HVS
1995b
Материнская агрессия у красных болотных раков ( Procambarus clarkii , Girard): связь между репродуктивным статусом и исходом агрессивных встреч с самцами и самками сородичи.
Поведение
132
107
— 125
Фиглер
M. H.
, Blank
G. S.
и Peeke
H. V. S.
2001
Материнская территориальность как стратегия защиты потомства у красных болотных раков Procambarus clarkii , Girard.
Агрессивное поведение
27
391
— 403
Фиглер
MH
, Blank
GS
и Peeke
HVS
2005
HVS
2005
Resident Shelter Procambarus clarkii Жирар и другие злоумышленники, различающиеся по полу и репродуктивному статусу.
Поведение и физиология в морской и пресноводной среде
38
237
— 248
Galeotti
P.
, Pupin
F.
, Rubolini
D.
, Sacchi
, Sacchi
, Nardi
PA
и Fasola
M.
2007
Влияние статуса спаривания на поведение при копуляции и расход спермы у пресноводных раков Austropotamobius italicus .
Поведенческая экология и социобиология
61
711
— 718
Галеотти
P.
, Rubolini
D.
, Fea
G.
, Ghia
Nardi
P.
, Gherardi
F.
и Fasola
M.
2006
Самки пресноводных раков корректируют размер яиц и кладок в зависимости от нескольких признаков самцов.
Труды Лондонского королевского общества (B)
273
1105
— 1110
Griffith
S.
и Sheldon
B.
2001
Фенотипическая пластичность в выражении сексуально отобранных черт: игнорируемые компоненты вариации.
Поведение животных
61
987
— 993
Guiasu
R. C.
и Dunham
D.W.
1998
Межформные агонистические соревнования у самцов раков, Cambarus robustus (Decapoda, Cambaridae).
Биология беспозвоночных
117
144
— 154
Hagen
HA
1870
Иллюстрированный каталог Музея сравнительной зоологии Гарвардского колледжа
2
11 viii + 109 страниц
Hamr
P.
2002
Orconectes .
585
— 608
Holdich
DM
Биология пресноводных раков
Blackwell Science
Oxford
Hays
WL
1994
Statistics
Harcourt
Jennions
M.
и Petrie
M.
2000
Почему самки размножаются? Обзор генетических преимуществ.
Биологические обзоры
75
21
— 64
Johnsson
J. I.
1997
Индивидуальное распознавание влияет на агрессию и отношения доминирования у радужной форели, Oncorhynchus mykiss .
Этология
103
267
— 282
Кирк
RE
1995
Экспериментальный план: процедуры для поведенческих наук
Brooks / Cole Publishing Company
2 Pacific Grove4,
, Pacific Grove4,
,
, Pacific Grove4,
Клокер
С.A.
и Strayer
D. L.
2004
Взаимодействия среди инвазивных раков ( Orconectes rusticus ), местных раков ( Orconectes limosus ) и местных двустворчатых моллюсков (Sphaeriidae и Unionidae).
Северо-восточный натуралист
11
167
— 178
Кодрик-Браун
A.
1992
Доминирование самцов может повысить успешность спаривания у гуппи.
Поведение животных
44
165
— 167
Кокко
H.
, Brooks
R.
, Jennions
M.
и Morley
J.
2002 Эволюция выбора партнера и предубеждений в отношении спаривания.
Труды Лондонского королевского общества (B)
270
653
— 664
Мэтьюз
Л. М.
и Warren
A. H.
2008
Новый рак из рода Orconectes Cope, 1872 г. из южной части Новой Англии (Crustacea: Decapoda: Cambariidae).
Труды Вашингтонского биологического общества
121
374
— 381
Мейнард Смит
Дж.
1978
Теория оптимизации в эволюции.
Ежегодный обзор экологии и систематики
9
31
— 56
Мур
P.A.
2007
Агонистическое поведение пресноводных раков — влияние внутренних и внешних факторов на агрессивные встречи и доминирование.
90
— 114
Даффи
EJ
и Тиль
M.
Эволюционная экология социальных и половых систем — ракообразные как модельные организмы
Harvard University Press
New York
Moore
PA
и Bergman
D.A.
2005
Запах успеха и неудачи: роль внутренних и внешних химических сигналов в социальном поведении раков.
Интегративная и сравнительная биология
45
650
— 657
Pavey
CR
и Fielder
DR
1996
Влияние разницы в размерах на агонистическое поведение пресноводных раков Cherax cuspidatas (Decapoda: Parastacidae).
Journal of Zoology
238
445
— 457
Pecor
K. W.
2006
Компромисс между преследованием и предотвращением хищничества у зрелых раков, Orconectes virilis .
Поведение и физиология в морской и пресноводной среде
39
229
— 233
Qvarnstrom
A.
1997
Экспериментально увеличенный размер значка увеличивает конкуренцию мужчин и снижает заботу родителей о самцах в ошейниковой мухоловке.
Протоколы Лондонского королевского общества (B)
264
1225
— 1231
Qvarnstrom
A.
и Forsgren
E.
1998
Если женщины предпочитают доминирующих мужчин ?
Тенденции в экологии и эволюции
13
498
— 501
Роли
MJ
и McGuire
MT
1989
Влияние самок на доминирование самцов у 9000 обезьян в неволе7 Cercopithecus aethiops sabaeus .
Поведение животных
38
59
— 67
Reichard
M.
, Bryja
J.
, Ondračková
M.
, Dávidová
, P.
и Smith
C.
2005
Половой отбор на доминирование самца снижает возможности выбора самки у европейского горчака ( Rhodeus sericeus ).
Молекулярная экология
14
1533
— 1542
Рейнольдс
Дж. Д.
2002
Рост и воспроизводство.
152
— 191
Holdich
DM
Биология пресноводных раков
Blackwell Science
Oxford
Rubolini
D.
, Galeotti
P.
, Ferrari
, Спайрани
М.
, Bernini
F.
и Fasola
M.
2006
Распределение сперматозоидов в зависимости от мужских признаков, размера самки и поведения при совокуплении у пресноводных раков.
Поведенческая экология и социобиология
60
212
— 219
Schroeder
L.
и Huber
R.
2001
Стратегии борьбы различаются размером и аллометрическим ростом раки, Orconectes rusticus .
Поведение
138
1437
— 1449
Stocker
AM
и Huber
R.
2001
Стратегии борьбы с раками Orconectes rustidae состояние голода и наличие пищевых сигналов.
Этология
107
727
— 736
Tilson
R.H.
и Hamilton
W.J.
III 1984
Социальное доминирование и особенности питания пятнистых гиен.
Поведение животных
32
715
— 724
Trivers
R. L.
1972
Родительское вложение и половой отбор.
139
— 179
Campbell
B.
Половой отбор и происхождение человека
Aldine
Chicago
Walker
D.
, Porter
B.A.
и Avise
J. C.
2002
Оценка генетического родства у раков Orconectes placidus , беспозвоночное с высокой плодовитостью, нуждающееся в расширенном материнском уходе.
Молекулярная экология
11
2115
— 2122
Wetzel
J.
2002
Изменение формы взрослых самок раков рода Orconectes (Decapoda: Cambaridae).
American Midland Naturalist
147
326
— 337
Winer
BJ
, Brown
DR
и Michels
KM
1991
Принципы статистического проектирования
McGraw-Hill
New York
Zulandt-Schneider
RA
, Huber
R.
и Moore
P.A.
2001
Распознавание индивидуума и статуса у раков, Orconectes rusticus : влияние выделения мочи на динамику схватки.
Поведение
138
137
— 153
© The Crustacean Society, 2016. Опубликовано Brill NV, Leiden
Раки-мутанты, полностью самки, размножающиеся путем клонирования
Марморреба сейчас настолько распространены на Мадагаскаре, что они угрожают местным видам.Андре Карват / CC BY-SA 2.5 Мраморные раки очень похожи на любых других пресноводных ракообразных. У него два когтя, десять ног и красивый сине-коричневый мраморный панцирь. Однако это шестидюймовое существо, обитающее в ручьях и озерах по всему миру, представляет гораздо более зловещие возможности, чем вы могли ожидать. Его новое научное название дает несколько подсказок: Procambarus virginalis. Все мраморные раки — самки, и они размножаются путем клонирования самих себя.
Фрэнк Лыко, биолог из Немецкого центра исследования рака, впервые услышал о мраморных раках от владельца аквариума, который в 1995 году подобрал «техасских раков» в зоомагазине.Они были поразительно большими и откладывали огромные партии яиц — сотни за один присест. Вскоре, как сообщает New York Times , любитель был осажден таким количеством раков, что раздавал их своим друзьям. И вскоре после этого marmorkrebs, как их называют по-немецки, стали появляться в зоомагазинах в Европе.
В этих раках было что-то очень странное. Все они были самками, и все они отложили сотни яиц без спаривания. Из этих яиц, в свою очередь, вылупились еще сотни самок, каждая из которых вырастает полностью способной к воспроизводству.В 2003 году ученые секвенировали их ДНК и подтвердили то, что многие владельцы уже считали так: каждый детеныш раков был клоном своей матери, и они наполняли рыбные ловушки Европы с угрожающей скоростью. «Люди начинают с одного животного, а через год у них будет пара сотен», — сказал Лико в интервью Times .
Всего 25 лет назад мраморных раков вообще не существовало. Теперь их можно найти в дикой природе миллионами в Германии, Чехии, Венгрии, Хорватии, Украине, Японии и Мадагаскаре.
Онлайн-совет для владельцев марморкребов ясен: они размножаются, как кролики. «Хотя 10 галлонов [аквариума] хватит на пару месяцев, — предупреждает один сайт, — эти 10 галлонов будут становиться все меньше и меньше по мере размножения этих животных. … Хорошее практическое правило: чем больше колония, тем больше аквариум. Начните с 40 галлонов и увеличьте объем, если можете себе это позволить ». Любители открыли эту истину на свой счет, что побудило многих выбрасывать своих дополнительных домашних животных в озера и ручьи.
В любых условиях раки прекрасно себя чувствуют.Оказывается, Marmorkrebs настолько же выносливы, насколько и плодовиты. А когда в водоеме их слишком много, они просто перемещаются. Times сообщает, что они пройдут сотни ярдов, чтобы найти новые озера или ручьи, где один экземпляр может произвести целую популяцию.
За последние пять лет Лико и его коллеги секвенировали геном мраморных раков, опубликовав свои результаты на этой неделе в журнале Nature Ecology & Evolution .Похоже, что эти раки являются результатом единственной радикальной мутации у речного рака, обитающего в реке Сатилла во Флориде и Джорджии. Около трех десятилетий назад спарились два плоских рака. При нормальном воспроизводстве каждая половая клетка имеет по одной копии каждой хромосомы. Но с одним из этих двух первых раков было что-то не так — мутация, которая оставила у него две копии в его половой клетке. Почему-то потомство оказалось выносливым и в отличном состоянии. Невероятно, но у него было по три копии каждой хромосомы и способность к бесполому размножению, не требуя участия мужчины.
Итак, захватит ли Землю армия клонов самок мраморных раков? «Это зависит от обстоятельств», — сказал Лыко « Times ». «Может быть, они просто проживут 100 000 лет. Лично для меня это займет много времени, но в процессе эволюции это будет просто отметкой на радаре ». По его словам, у их бесполого размножения есть свои преимущества и недостатки. Рост популяции стремительный, один экземпляр способен произвести сотни плодовитых потомков. Но они тоже уязвимы. При отсутствии генетических вариаций, если болезнь может уничтожить один клон, все остальные с большой вероятностью будут уничтожены.Однако сколько времени потребуется, чтобы появился такой патоген, остается загадкой. А пока во всем мире раки чувствуют себя как дома.
ADW: Cambarus bartonii: ИНФОРМАЦИЯ
Географический диапазон
Cambarus bartonii обитает в восточной и южной частях Соединенных Штатов, а также в юго-восточной части Канады. Однако чаще всего они встречаются в диапазоне от Нью-Брансуика, Канада, до северной Джорджии и в восточных частях Кентукки и Теннесси.(Зеленый, 1967)
Среда обитания
Cambarus bartonii обитают на дне ручьев, ручьев, малых рек и озер. Они строят норы, иногда называемые «дымоходами». Их норы могут быть простыми дуплами под камнем или более замысловатыми, с боковыми проходами. Дымоходы расположены вдоль кромки воды. Большая часть конструкции находится под водой, но вершина выступает и напоминает дымоход. Дымоходы различаются по размеру, самое большое отверстие составляет около восьми сантиметров.(Макман, 1960)
- озера и пруды
- реки и ручьи
Физическое описание
Cambarus bartonii — пресноводное ракообразное. При первом осмотре он выглядит «как омар». У него острая морда, а глаза на подвижных стеблях. Тонкий, но прочный экзоскелет темно-коричневого цвета, иногда со слегка красным оттенком.Точный оттенок зависит от того, как выглядит нижний субстрат среды обитания.
Из придатков наиболее заметны хелипеды. Эти придатки прикрепляются к грудной клетке и также называются первыми ходовыми ногами. Однако у этих придатков есть когти, и они используются для защиты и ловли пищи, а не для ходьбы. Cambarus bartonii также имеет четыре другие пары ходильных ног, прикрепленных к грудной клетке. Брюшные придатки Cambarus bartonii называются пловцами или плеоподами, они намного меньше ходячих ног и не подходят для плавания.Мужские пловцы модифицированы для передачи пакетов спермы самкам во время размножения и имеют лопатчатую форму. То, что похоже на веер на конце брюшка, на самом деле представляет собой множество широких плоских придатков, называемых уроподами. Все эти придатки прикреплены к брюшной стороне Cambarus bartonii , и все они двуветвистые. В целом живот большой и обычно расширенный; однако он может быть согнут под головогрудью. Первый сегмент брюшной полости обычно меньше, чем кзади от него.
Cambarus bartonii имеет открытую систему кровообращения. У него ромбовидное сердце, расположенное кпереди от брюшных сегментов по средней линии спины. Сердце окружено тонким перикардиальным мешком. Прямо перед сердцем лежат гонады. Семенники белые, а яичники оранжевые. Вдоль дорсальной средней линии головогруди лежат сердечный желудок и пилорический желудок. Сразу кзади и сбоку от желудка расположена большая пищеварительная железа.Брюшной нервный тяж лежит под внутренними органами, а мозг находится между стебельками глаз и под ними. (Рассел-Хантер, 1979; Водопич, Мур, 1999)
Репродукция
Спаривание чаще всего происходит весной, но может происходить и летом. Спаривание обычно происходит ночью, потому что шансы встретить друг друга самцом и самкой ночью в девять раз выше.
Размножение предполагает спаривание и может происходить двумя способами.Первый — это отложение спермы в семяприемник у самки. Это происходит, когда сперматозоид самца течет по бороздкам первых плеоподов и попадает в приемник самки. Сперма выходит из самца раков у основания пятой пары ходильных ног через пору. Яйца выпускаются у основания третьей пары ходильных ног. Другая форма размножения включает перенос сперматофора, и в этом случае оплодотворение является внутренним.
В любом случае оплодотворенные яйца сохраняются для созревания на плеоподах самки.Они вылупляются на плеоподах и остаются привязанными к матери вскоре после второй линьки. Говорят, что самка, несущая яйца, находится «в ягоде», потому что масса яиц похожа на ягоду. Самки чаще всего «в ягоде» в мае и июне. (Робертс, 1994; Водопич, Мур, 1999)
Поведение
Cambarus bartonii медленно ходите по дну прудов, ручьев или ручьев в поисках пищи. В случае угрозы они могут стрелять назад, чтобы быстро избежать опасности.Они совершают это быстрое движение назад, сокращая мощные мышцы живота. Cambarus bartonii используют хелипедов для защиты и поиска пищи. Хищники на C. bartonii включают различные виды птиц и черепах. Cambarus bartonii наиболее активен ночью, потому что именно тогда он питается. (Buchsbaum и др., 1987)
Привычки питания
Cambarus bartonii — хищник и падальщик.Он питается разлагающимися органическими остатками, но также ловит мелких животных. Его основными источниками пищи являются улитки, водоросли, личинки насекомых, различные виды червей и головастиков. Он находит пищу на дне источника, в котором обитает, или в почве рядом с водой. (Банистер и Кэмпбелл, 1985)
- амфибии
- яйца
- насекомые
- моллюски
- водные или морские черви
- водные ракообразные
Экономическое значение для людей: положительный результат
Раки — огромная индустрия для многих стран, особенно для Франции и США.Ежегодно потребляется от 400 000 до 800 000 кг (от 880 000 до 1 960 000 фунтов). Ежегодно в искусственных водоемах выращивают 2,6 миллиона фунтов раков. Это создает много рабочих мест для людей, занятых в рыбной отрасли.
Раки полезны и являются источником белка. Сейчас их едят в Соединенных Штатах и во многих странах мира.
Раки также могут иметь лечебные свойства. В Кении — микроскопический кровяной сосальщик.шистом вызывает инфекцию у многих людей. Шистом проникает через кожу и перемещается к мочевому пузырю и другим основным органам, вызывая серьезные повреждения. Ученые выяснили, что личинки шистома вылупляются в пресноводных улитках. Они также обнаружили, что у раков есть аппетит к этим пресноводным улиткам. Размещение большого количества раков (хотя не говорится, какие именно виды будут использоваться; у большинства видов есть аппетит к улиткам) в этих озерах может уменьшить популяцию улиток и, в свою очередь, снизить количество инфекций.
Cambarus bartonii закапывает почву вдоль ручьев, прудов и рек. Это помогает в сельском хозяйстве, потому что перекопка обогащает почву питательными веществами. На многих рисовых фермах раки являются второй культурой, потому что фермеры привозят их после уборки риса, чтобы помочь почве получить много питательных веществ. Затем фермеры собирают раков, когда снова наступает сезон риса. Это не является преимуществом для конкретного вида, потому что в мире существует множество рисовых ферм.
Раки также используются для мониторинга экологического состояния ручьев и рек. В частности, их можно проанализировать на предмет наличия в их тканях определенных загрязнителей. («Раки», 1998; Банистер и Кэмпбелл, 1985)
Экономическое значение для людей: отрицательное значение
Нет никаких известных побочных эффектов Cambarus bartonii на человека.
Статус сохранения
Известно, что этот вид не находится под угрозой исчезновения.
Авторы
Стефани Дженкинсон (автор), Юго-Западный университет, Стефани Фабрициус (редактор), Юго-Западный университет.
Глоссарий
- Неарктика
человек проживает в Неарктической биогеографической провинции, северной части Нового Света. Это включает Гренландию, канадские арктические острова и всю Северную Америку вплоть до юга до высокогорья центральной Мексики.
- двусторонняя симметрия
, имеющий такую симметрию тела, что животное можно разделить в одной плоскости на две зеркальные половины. У животных с двусторонней симметрией есть спинная и вентральная стороны, а также передний и задний концы. Синапоморфия билатериев.
- детрит
частицы органического материала от мертвых и разлагающихся организмов. Детрит — результат деятельности деструкторов (организмов, разлагающих органический материал).
- экзотермический
животных, которые должны использовать тепло, полученное из окружающей среды, и поведенческие адаптации для регулирования температуры тела
- пресная вода
в основном обитает в несоленой воде.
- гетеротермический
имеет температуру тела, которая колеблется в зависимости от температуры окружающей среды; не имеющий механизма или плохо развитый механизм регулирования внутренней температуры тела.
- гибернация
состояние, в которое некоторые животные заходят зимой, когда нормальные физиологические процессы значительно замедляются, что снижает потребность животных в энергии.Акт или условие прохождения зимы в оцепенении или состоянии покоя, обычно предполагающее отказ от гомойотермии у млекопитающих.
- итеропарное
потомков производятся более чем в одной группе (пометы, кладки и т. Д.) И в течение нескольких сезонов (или других периодов, благоприятных для воспроизводства). По определению, итеропородящие животные должны выживать в течение нескольких сезонов (или периодических изменений состояния).
- подвижный
, имеющий возможность перемещаться с одного места на другое.
- натуральный
для плавания
- родной диапазон
район, в котором животное встречается в природе, регион, в котором оно является эндемиком.
- ночной
активен ночью
- всеядное
животное, которое в основном ест всевозможные продукты, включая растения и животных
- яйцекладущие
размножение, при котором яйца выпускает самка; развитие потомства происходит вне организма матери.
- сезонное разведение
разведение привязано к определенному сезону
- малоподвижный
остается на том же участке
- половой
воспроизводство, включающее сочетание генетического вклада двух особей, мужчины и женщины
- наземный
Проживает на земле.
Список литературы
1998. Раки. Всемирная книжная энциклопедия. Лондон: World Book Inc ..
Банистер, К., А. Кэмпбелл. 1985. Омары и пресноводные раки. Стр. 236-239 в Энциклопедии водной жизни. Нью-Йорк: Equinox Ltd.
Buchsbaum, R., M. Buchsbaum, J. Pearse, V. Pearse. 1987. Животные без позвоночника. Чикаго: Издательство Чикагского университета.
Грин, Дж. 1967. Ракообразные. Стр. 172-175 в Энциклопедии жизни животных Ларусс. Нью-Йорк: Книжная компания Mc Graw-Hill.
Макман Л. 1960. Случай строительства дымохода раками * Cambarus bartonii *. Экология, 41: 383-385.
Робертс, Т. 1994. Свет, химические вещества для глаз и некоторые другие факторы как регуляторы общественной активности раков. Экологические монографии, 14: с. 376-378.
Рассел-Хантер, В.1979. Жизнь беспозвоночных. Нью-Йорк: McMillan Publishing Co Inc.
Водопич Д., Мур Р. 1999. Руководство лаборатории биологии. Берр Ридж: У. Х. Фриман и компания.
Раки (от NC WINS) | NCpedia
Камбарус бартони Написано Сарой Пятница
Обновлено Робертом Николсом
Комиссия по ресурсам дикой природы Северной Каролины.
Классификация Класс: Ракообразные
Отряд: Декапода «десятиногая»
Средний размер Около 3 дюймовв длину
Еда Питаются в основном водной растительностью, но также едят червей,
улиток, головастиков и других мелких живых или мертвых существ.
Разведение Большинство дает молодь весной. Самка откладывает сотни яиц; вылупление около 85%. Количество выводков разное. Ученые не уверены, сколько времени требуется для вылупления яиц, но по их оценкам, месяц или больше. Когда яйца вылупляются, появляются крошечные прозрачные версии взрослых раков размером около 1/6 дюйма.долго.
Молодые Молодь цепляется за самку через три линьки, пока не достигнет размера около 1 1/2 дюйма. Раки не проходят личиночную стадию.
Ожидаемая продолжительность жизни Примерно 3-5 лет.
Диапазон и распространение
Некоторые раки являются универсальными и встречаются на большей части территории Северной Каролины, но большинство из них ограничено небольшими географическими районами. Раки-роющие Гринсборо, Cambarus catagius, встречаются только в районе Гринсборо, а колючие раки Северной Каролины, Orconectes carolinensis, встречаются только в бассейнах рек Тар и Нойз на прибрежной равнине.
Общая информация Летом переверните скалу почти в любом ручье в Северной Каролине, и велика вероятность, что рак отскочит от вас назад или даже защитит скалу своими когтями, направленными на вас. Существует по крайней мере 40 различных видов этого обычного пресноводного ракообразного, которые обитают в ручьях, прудах и норах Тар Хил от гор до побережья. Многие из этих видов не встречаются больше нигде на Земле.
Описание Раки выглядят как миниатюрные омары с передней парой сильных щипающих когтей, бронированным телом и широким хвостом.Как и у лобстеров, у раков есть 3 основные части тела: голова, грудная клетка и живот, а также твердый экзоскелет, который защищает их мягкие ткани и органы. Передняя часть тела жесткая, но задняя часть, брюшко или хвост, имеет подвижные сегменты. В области головы 2 пары антенн и крошечные глазки помогают ракам воспринимать окружающую среду. В грудной клетке у раков по 5 ног с каждой стороны (тела) и мягкие перистые жабры, которые используются для дыхания. Первая пара ног — это большие когти или клешни, которые помогают ракам ловить и удерживать добычу.Следующие 4 пары ног — это ходячие ноги; первые 2 пары — это маленькие клешни, используемые для обработки пищи. Под хвостом и после последней пары ходильных ног есть более мелкие и более тонкие придатки, похожие на ноги, называемые пловцами, которые помогают плавать, переносить яйца и удерживать молодых раков. В зависимости от вида окраска раков варьируется от малиново-красного до кобальтово-синего. Некоторые из них могут быть розовыми, коричневыми, зеленовато-черными или даже аквамариновыми.
История и статус Наши биологи ежегодно открывают и описывают новые виды раков, а также документируют распространение экзотических раков в штате.По мере завершения новых исследований биологи продолжают документировать расширение ареала, а также сокращение некоторых раков Северной Каролины.
Среда обитания и привычки
Раки обитают в ручьях, прудах, озерах и болотах по всей Северной Каролине. Обитатели ручьев предпочитают быстро движущиеся и насыщенные кислородом реки и ручьи гор и предгорий. В более медленных ручьях, например на Прибрежной равнине, раки прячутся под камнями и бревнами для защиты.Раков также можно встретить в прудах, озерах и в стоячей воде в придорожных канавах. Некоторые раки зарываются в землю и могут рыть разветвляющиеся туннели глубиной до 15 футов в илистых заболоченных местах. Большинство нор в среднем составляют 2–3 фута в глубину, и их можно найти на приличном расстоянии от стоячей воды. Раки-роющие, обитающие в основном на Прибрежной равнине и в восточной части Северной Каролины, во время копания часто выбрасывают глиняные «дымоходы».
Раки становятся наиболее активными с наступлением темноты или на рассвете и выходят из своих укрытий, чтобы покормиться.Эти стойкие ракообразные питаются в основном водной растительностью, но они также могут есть мелких, живых или мертвых животных, таких как черви, улитки, головастики и рыба. Раки часто становятся добычей других животных, таких как лягушки, черепахи, окуни, еноты, болотные птицы и ястребы. Но раки — бойцы. Лучшая защита — мощный хвост, который отбрасывает раков назад. Если они теряют коготь или ногу в бою, на их месте вырастает новый.
Большинство видов раков в Северной Каролине дают потомство ранней весной, когда много еды и дождя.Некоторые виды размножаются зимой. Для спаривания самец рака хватает самку и переворачивает ее так, чтобы брюшная сторона находилась вместе. Затем самец передает сперму самке через свою первую пару плавников, называемых гоноподом. При оплодотворении сотни крошечных яиц прикрепляются к плавникам самки с липкими выделениями, называемыми яицами. Когда к самке раков прикрепляются оплодотворенные яйца, говорят, что она в ягоде. Более крупные самки обычно откладывают больше яиц.
В отличие от многих других ракообразных, раки не проходят личиночную стадию, а линяют около 6 раз в первые 7-8 месяцев жизни.Линька или выпадение панциря необходимо, потому что раки перерастают свою броню. Они продолжают линять более медленными темпами всю оставшуюся жизнь. Когда они линяют, старый скелет отпадает, и раки с мягким панцирем быстро растут, так как поглощают большое количество воды. Незащищенные, они прячутся после линьки и менее агрессивны, пока через несколько дней не затвердеет новый скелет. Раки прекрасно себя чувствуют в незагрязненной воде, и их количество является хорошим признаком хорошей экологии.
Взаимодействие с людьми Раки ценятся не только с экологической точки зрения, но и с экономической точки зрения.Более крупные раки, такие как красные болотные раки, считаются деликатесом каджунской кухни. Раки также используются в качестве приманки для рыбной ловли, и это привело к некоторым случайным проникновениям неместных раков в новые районы. Неместные раки вредят нашей экосистеме, конкурируя с местными видами тархилов и уничтожая водные растения, которые служат пищей и убежищем для местных животных. ПОЖАЛУЙСТА, НЕ выпускайте раков в водные пути Северной Каролины !!! Попав сюда, их невозможно искоренить! Вот совет для тех, кому не посчастливилось попасть не на тот конец раков.У них есть острый укус, но он укусил их только в качестве последней попытки самообороны. Если им дадут шанс сбежать, они это сделают. Так что, если вы когда-нибудь обнаружите, что к вам прицепились раки, просто
воткните пораженную часть анатомии обратно в воду, где раки будут чувствовать себя в большей безопасности и уплывут. Этот метод также работает с черепахами.
Примечание: «раки» относятся к одному виду или отдельному экземпляру, тогда как «раки» относятся ко многим видам.
Судьба золотистых желез и нерестовое поведение у неместных раков: сравнение гонохорических речных раков и партеногенетических мраморных раков.
Abstract
В период до нереста у самок пресноводных раков развиваются золотистые железы на нижней стороне плеона.Эти железы производят слизь для временной структуры, похожей на палатку, в которой яйца оплодотворяются и прикрепляются к плеоподам. Долгосрочное наблюдение за самками рака Procambarus fallax , размножающихся двуполым путем, содержащихся в неволе, показало, что голые железы развиваются в конце зимы и в конце лета каждого года независимо от присутствия самцов. Однако, в отличие от спарившихся самок, самки без спаривания не образуют ни оплодотворяющей палатки, ни нереста. Их голые железы сохранялись необычно долго и исчезли только во время следующей линьки.Судя по всему, женщины используют информацию о наличии сперматозоидов либо для воспроизводства, либо для сохранения ресурсов. Мраморный рак, Procambarus virginalis , партеногенетический потомок слепого рака, развил голые железы примерно в одни и те же периоды года, но нерестился, несмотря на отсутствие спаривания. Эти находки указывают на то, что на пути от гонохоризма к партеногенезу регуляция активности голых желез и нереста была отделена от спаривания. Таким образом, пара видов Procambarus fallax / Procambarus virginalis кажется особенно подходящей для исследования физиологических, молекулярных и генетических механизмов, лежащих в основе нереста пресноводных раков.
Введение
За несколько недель до нереста у пресноводных раков на нижней стороне плеона развиваются выступающие белесые блестящие железы (Reynolds 2002). Эти железы сначала выглядят как слабые кремово-беловатые пятна и становятся все более и более отчетливыми, что является хорошим индикатором предстоящего нереста (Jussila et al. 1996). Незадолго до откладывания яиц голые железы выделяют полупрозрачную слизь, которая образует тентовую структуру на нижней стороне грудной клетки и плеона (Andrews 1904; Vogt 2016).В этой студенистой массе сперматозоиды мобилизуются из сперматофоров, которые прикреплены или хранятся внутри, в зависимости от семейства раков (Reynolds 2002; Vogt 2002; Niksirat et al. 2013). Палатка для оплодотворения дополнительно обеспечивает прикрепление оплодотворенных яиц к плеоподам, где они высиживаются в течение нескольких недель до вылупления первой молоди и далее (Vogt and Tolley 2004). Глэровые железы и палатка оплодотворения являются уникальными особенностями пресноводных раков и встречаются во всех трех семействах: Astacidae, Cambaridae и Parastacidae (Andrews 1906; Mason 1970; Thomas and Crawley 1975; Jussila et al.1996; Фогт 2016). Более простые цементные железы, которые только способствуют прикреплению яиц к плеоподам, встречаются и у других Decapoda (Adiyodi and Anilkumar 1988).
Во время конференций по ракообразным и студенческих курсов я часто сталкивался с вопросом, что происходит с желчными железами и зрелыми ооцитами, если самки раков не спариваются. К сожалению, нет опубликованных данных о судьбе желчных желез и очень мало данных о нересте у несвязанных самок (Woodlock, Reynolds 1988; Reynolds et al.1992; Холдич и др. 1995; Tropea et al. 2010; Buřič et al. 2011). Одна из возможностей состоит в том, что несвязанные самки нерестятся как спарившиеся самки, но неоплодотворенные яйца разлагаются. В качестве альтернативы слизь и зрелые ооциты могут реабсорбироваться, чтобы избежать растраты ресурсов. В настоящем сообщении я сообщаю о судьбе головых желез и нерестовом поведении у выращенных в лаборатории спарившихся и не спарившихся речных раков Procambarus fallax и их партеногенетически воспроизводящегося родственника, мраморного рака Procambarus virginalis .У мраморных раков нет самцов, и они произошли от речных раков в результате аутотриплоидизации и сопутствующего перехода от гонохоризма к партеногенезу (Scholtz et al. 2003; Martin et al. 2010; 2016; Vogt et al. 2015). Недавно они были признаны отдельным видом из-за репродуктивной и географической изоляции от Procambarus fallax , уникальных генетических и эпигенетических признаков, а также превосходного роста и плодовитости (Vogt et al. 2015).
Материалы и методы
Самки речного рака Procambarus fallax (Hagen 1870) (Decapoda: Cambaridae) были отделены от своих сородичей-самцов на пятой стадии молоди, первой стадии жизни, которая позволяет определить пол по внешнему признаку.В последующие пять месяцев их держали вместе в женской группе, а затем по отдельности. Три из этих самок остались без спаривания, а еще три самки были повязаны с самцами в течение сезонов размножения весной 2015, осенью 2015 и весной 2016. Спаривание производилось путем спаривания отдельных самок с гладкими железами и отдельных самцов с крючками на седалищных костях 3-й и 3-й седалищных желез. 4-е переоподы (показатели половой зрелости) в контейнере 30 × 25 × 20 см. Копуляция считалась успешной, когда самцы переворачивали самок на спину и вставляли их копулятивные органы в приемник спермы (annulus ventralis) самки не менее чем на 30 минут.Все спарившиеся и не спарившиеся самки содержались индивидуально в пластиковых контейнерах 30 × 25 × 20 см, снабженных гравием и укрытием. В качестве источника воды использовалась водопроводная вода, которую меняли раз в неделю. Температура воды колебалась от ~ 15 ° C зимой до ~ 25 ° C летом, и фотопериод был естественным режимом окружающей среды Гейдельберга (Германия). Раков кормили ежедневно ad libitum гранулами TetraWafer Mix. Период наблюдений длился с 1 июля 2014 г. по 31 мая 2016 г.Я дал время в неделях только потому, что в начале созревания слепой железы трудно решить, началось ли уже развитие или нет. Данные по Procambarus fallax были сопоставлены с моими записями о наличии блестящих желез у партеногенетических мраморных раков Procambarus virginalis , которые я собирал за последние 13 лет. Мраморных раков выращивали в тех же условиях, что и речных раков, индивидуально или коллективно.
Результаты
У самок обоих видов раков в период размножения развиваются голые железы. Эти железы были расположены в последней грудной части грудины, грудины и плевре плеона, уропод и плеопод (рис. 1А, В). Головые железы развивались автономно, в отсутствие самцов.
Рис. 1. Глеровские железы и приемник для сперматозоидов у раков. (A) Зрелые железы голени на нижней стороне плеона партеногенетической самки Procambarus virginalis .Беловатые железы расположены в грудины (стрелка), плевре (наконечник стрелки), уроподах (u) и плеоподах (p). Пруток: 5 мм. (B) Плеон самки Procambarus fallax без спаривания с обнаженными железами через 12 недель после их первого появления. Пруток: 2 мм. (C) Сканирующая электронная микрофотография брюшного кольца самки Procambarus virginalis . Бар: 200 мкм.
В трех проанализированных периодах воспроизводства нерестовые самки Procambarus fallax не спаривались, хотя у них регулярно развивались голые железы (Таблица 1).Напротив, спарившиеся самки обычно откладывали яйца. В последнем случае блестящие железы были видны в течение трех-пяти недель. Их опорожняли незадолго до кладки яиц, чтобы сформировать палатку для оплодотворения. У несвязанных самок золотистые железы сохранялись до 12 недель (таблица 1) и исчезли только во время следующего шелушения.
Таблица 1. Судьба золотистых желез и нерестовое поведение у не спарившихся и не спарившихся Procambarus fallax самок.
Мраморных раков, выращиваемых в лаборатории, в основном воспроизводятся два раза в год, как и речных раков (Vogt 2015).Некоторые особи размножаются только один-три раза в год. За 3-6 недель до нереста у них были обнаружены железы с увеличивающейся интенсивностью окрашивания. В отличие от несвязанных самок речного рака, партеногенетические самки мраморных раков производили палатки для оплодотворения и нерестились. Только в одном из 103 задокументированных случаев самка с голыми железами пропустила нерест и вместо этого выполнила линьку. Золотистые железы этого экземпляра исчезли во время линьки, как и у несвязанных раков.
Обсуждение
Сравнение не спарившихся самок гладкоплодных раков и партеногенетических мраморных раков выявило интересные различия в отношении нереста и судьбы гладких желез.Спаривающиеся самки и самки мраморных раков обычно нерестятся дважды в год (Vogt 2015). Эти нереста связаны с опорожнением желёз. У слабых раков оба события зависят от спаривания. Несвязанные Procambarus fallax самок не порождают и не опорожняют свои голые железы. Вместо этого блестящие железы сохраняются еще много недель, вероятно, в ожидании более позднего шанса спаривания. Это контрастирует с мраморными раками, у которых развиваются голые железы и нерестятся в отсутствие самцов.Очевидно, что информационная система, позволяющая различать условия спаривания и отсутствия спаривания, была потеряна или изменена во время перехода от гонохоризма к партеногенезу в паре видов Procambarus fallax / Procambarus virginalis .
Полное подавление нереста наблюдалось также у самок астцид Austropotamobius pallipes , Astacus leptodactylus и Pacifastacus leniusculus по данным Holdich et al.(1995). Напротив, Вудлок и Рейнольдс (1988) наблюдали нормальный нерест, но распад прикрепленных яиц у несвязанных самок Austropotamobius pallipes , которые содержались в парах самок. У самок выращенных в лабораторных условиях Cherax destructor (Parastacidae), не имеющих спаривания, нерестовое поведение было весьма разнообразным: около 20% нерестились при выращивании при 30 ° C, но не появлялись при 27 ° C (Tropea et al. 2010). Нерест не спарившихся самок произошел и у факультативно партеногенетических самок камбарид Orconectes limosus (Buřič et al.2011, 2013). Из 30 не спаривавшихся партеногенетических самок 28 нерестились, и только одна самка не отнерестилась (оставшаяся особь погибла) (Buřič et al. 2011), как и у мраморных раков. Причины различий в нерестовом поведении самок, не имеющих спаривания, между указанными размножающимися половым путем видами и внутри них неизвестны, но могут зависеть от видовых различий, условий окружающей среды, предэкспериментального опыта и статуса питания особей. Информация о судьбе желёзок отсутствует во всех цитированных выше сообщениях.Подавление секреции и нереста гололеда можно интерпретировать как энергосберегающую стратегию.
Созревание ооцитов не зависит от присутствия самцов, как показали все вышеупомянутые исследования. То же самое касается созревания желёзок, которые развиваются за несколько недель до нереста. Оба процесса, по-видимому, тесно связаны и вызываются факторами окружающей среды, в основном температурой и световым периодом (Reynolds 2002; Vogt 2015), которые влияют на гормональную систему. Созревание яичников ингибируется гормоном, ингибирующим вителлогенез (VIH) из системы X-орган-синусовая железа, и стимулируется гормоном, стимулирующим яичники (OSH), из ганглиев грудных нервов (Reynolds 2002; Vogt 2002).Созревание блестящих желез, вероятно, регулируется теми же гормонами.
Спаривание может быть одним из спусковых механизмов для нереста, как предполагает Рейнольдс (2002). Это предположение подтверждается экспериментами Holdich et al. (1995), которые спарили самок с самцами другого вида. Такие самки обычно нерестились, но икринки не развивались. Однако в естественных условиях между спариванием и нерестом часто бывает много дней или даже недель, что позволяет предположить, что нерест также может зависеть от воспоминаний о спаривании самкой или ощущения присутствия спермы.У астацидных и парастакидных раков сперматофоры прикреплены снаружи к грудным пластинам грудины и хвостовому вееру (Reynolds 2002; Vogt 2002; Niksirat et al.2013), и, таким образом, самки могут легко ощущать присутствие сперматозоидов своими переоподами. У Cambaridae сперма хранится внутри вентрального кольца (рис. 1C) (Reynolds 2002; Vogt et al. 2004), и в этом случае механорецепторы могут дать информацию о состоянии ее наполнения. №
Неизвестно, могут ли раки добровольно опорожнять железы и яичники за счет сокращения мускулатуры желез и яичников.Более вероятно, что оба процесса являются непроизвольными и регулируются гормонами, такими как простагландин F 2α , который, как было показано, резко усиливается на заключительной стадии оогенеза у Procambarus paeninsulanus (Spaziani et al. 1995). Тесты in vitro показали, что этот гормон вызывает сокращение ткани яичников в зависимости от дозы.
Настоящее исследование является первым отчетом о судьбе железистой оболочки без спаривания раков, выращенных в контролируемых условиях.
Не было значительной разницы в общем размере самок между падениями | $ (n = 34, \, \ bar {x} = 35.203 {\ text {mm}}) $ | и пружина | $ (n = 28, \, \ bar {x} = 34.993 {\ text {mm}}, \, t = 0.219, \, {\ text {DF}} = 60, \, P = 0.827 ) $ | , либо для женщин в маленьком | $ ({\ text {fall:}} = 11, \, \ bar {x} = 30.572 {\ text {mm}} $ | ; | $ {\ текст {весна:}} n = 11, \, \ bar {x} = 31,046 {\ text {mm}}, \, t = -0,555, \, {\ text {DF}} = 20, \, P = 0,585 $ | ) или большой | $ ({\ text {fall:}} n = 23, \, \ bar {x} = 37,417 {\ text {mm}}; \, {\ text {spring:}} n = 17, \, \ bar {x} = 37,547 {\ text {mm}}, \, t = — 0,214, \, {\ text {DF}} \, = 38, \, P = 0.832) $ | размерные классы. Наблюдалась значительная разница в общем размере мужчин между падениями | $ (n = 34, \, \ bar {x} = 35.247 {\ text {mm}}) $ | и пружина | $ (n = 42, \, \ bar {x} = 33,136 {\ text {mm}}, \, t = 2,867, \, {\ text {DF}} = 74, \, P = 0,005 ) $ | , но не для маленьких | $ ({\ rm {fall}}: \, \, n = {\ rm {14}}, \, \, \ bar x = {\ rm {32} } .000 \, \, {\ rm {mm}}; \, \, {\ rm {spring}}: \, \, n = {\ rm {24}}, \, \, \ bar x = { \ rm {3}} 0. {\ rm {746}} \, \, {\ rm {mm}}, \, \, t = {\ rm {\ bar 2}}. {\ rm {376}} , \, \, {\ rm {DF}} = {\ rm {36}}, \, \, P = 0.0 {\ rm {23}}) $ | или большой | $ ({\ text {fall:}} n = 20, \, \ bar {x} = 37,520 {\ text {mm}}; \, {\ text {spring:}} n = 18, \ , \ bar {x} = 36,322 {\ text {mm}}, \, t = 2,275, \, {\ text {DF}} = 36, \, P = 0,029) $ | группы отдельно.
При сравнении времени, затраченного на общее агонистическое поведение, и лечение, и размер показали значительный эффект в различных сравнениях (таблица 2, рис. 1). Самцы | $ (n = 17) $ | и женщины | $ (n = 22) $ | в осеннем репродуктивном сезоне было обнаружено значительное влияние секса на общее время, затрачиваемое на агонистическое поведение, при этом самцы тратят больше времени на такое поведение, чем самки, и отсутствие значительного влияния размера на общее агонистическое поведение в этих двух группах.Существенной разницы между мужчинами | $ (n = 21) $ | и женщины | $ (n = 19) $ | летом, в период отсутствия репродукции, общее время, проведенное в агонистическом поведении, но и самцы, и самки для этого сравнения показали значительный эффект размера, при этом более крупные животные тратят значительно больше времени на агонистическое поведение, чем более мелкие животные. Сравнение репродуктивных и не репродуктивных особей не показало значительных различий в общем времени, проведенном в агонистическом поведении в зависимости от сезона или размера для любого пола, хотя для женщин взаимодействие между репродуктивным состоянием и размером имело значительный эффект и сохранялось в модель (но которая не имела значения при исправлении Бонферрони | $ {\ rm {\ alpha}} = 0.0125 $ | ).
Таблица 2 Результаты дисперсионного анализас фиксированными эффектами сезона, например, репродуктивные самцы по сравнению с непродуктивными самцами, или пол, например, репродуктивные самцы по сравнению с репродуктивными самками. Во всех сравнениях размер был включен как фактор; член взаимодействия был незначительным | $ (P> 0,05) $ | во всех случаях, кроме сравнения репродуктивных самок с непродуктивными самками, и был исключен из анализов, в которых он не был значимым.
| . | Фактор 1 (размер) . | Фактор 2 (сезон или пол) . | Фактор 1 * фактор2 . | |||
|---|---|---|---|---|---|---|
| Контрастность . | ф . | | $ P $ | . | ф . | | $ P $ | . | ф . | | $ P $ | . |
| Между сезонами: | ||||||
| 1.Репродуктивные самцы и непродуктивные самцы | 0,468 | 0,498 | 1,174 | 0,286 | — | — |
| 2. Репродуктивные самки и непродуктивные самки | 4,820 | 0,034 | 0,146 | 4,027 | 0,052 | |
| Между полами: | ||||||
| 3.Репродуктивные самцы и репродуктивные самки | 0,065 | 0,801 | 9,071 | 0,005 * | — | — |
| 4. Непродуктивные самцы и непродуктивные самки | 7,381 | 9 *0,025 | 0,876 | — | — | |
| . | Фактор 1 (размер) . | Фактор 2 (сезон или пол) . | Фактор 1 * фактор2 . | |||
|---|---|---|---|---|---|---|
| Контрастность . | ф . | | $ P $ | . | ф . | | $ P $ | . | ф . | | $ P $ | . |
| Между сезонами: | ||||||
| 1.Репродуктивные самцы и непродуктивные самцы | 0,468 | 0,498 | 1,174 | 0,286 | — | — |
| 2. Репродуктивные самки и непродуктивные самки | 4,820 | 0,034 | 0,146 | 4,027 | 0,052 | |
| Между полами: | ||||||
| 3.Репродуктивные самцы и репродуктивные самки | 0,065 | 0,801 | 9,071 | 0,005 * | — | — |
| 4. Непродуктивные самцы и непродуктивные самки | 7,381 | 9 *0,025 | 0,876 | — | — | |
Результаты ANOVA с фиксированными эффектами сезона, например, репродуктивные самцы по сравнению с непродуктивными самцами или полом, e.g, репродуктивные самцы по сравнению с репродуктивными самками. Во всех сравнениях размер был включен как фактор; член взаимодействия был незначительным | $ (P> 0,05) $ | во всех случаях, кроме сравнения репродуктивных самок с непродуктивными самками, и был исключен из анализов, в которых он не был значимым.
| . | Фактор 1 (размер) . | Фактор 2 (сезон или пол) . | Фактор 1 * фактор2 . | |||
|---|---|---|---|---|---|---|
| Контрастность . | ф . | | $ P $ | . | ф . | | $ P $ | . | ф . | | $ P $ | . |
| Между сезонами: | ||||||
| 1. Репродуктивные самцы по сравнению с непродуктивными самцами | 0.468 | 0,498 | 1,174 | 0,286 | — | — |
| 2. Репродуктивные самки по сравнению с непродуктивными самками | 4,820 | 0,034 | 2,207 | 0,146 | 4,0 | |
| Между полами: | ||||||
| 3. Репродуктивные самцы и репродуктивные самки | 0.065 | 0,801 | 9,071 | 0,005 * | — | — |
| 4. Непродуктивные самцы и непродуктивные самки | 7,381 | 0,010 * 76 | — | — | ||
| . | Фактор 1 (размер) . | Фактор 2 (сезон или пол) . | Фактор 1 * фактор2 . | |||
|---|---|---|---|---|---|---|
| Контрастность . | ф . | | $ P $ | . | ф . | | $ P $ | . | ф . | | $ P $ | . |
| Между сезонами: | ||||||
| 1.Репродуктивные самцы и непродуктивные самцы | 0,468 | 0,498 | 1,174 | 0,286 | — | — |
| 2. Репродуктивные самки и непродуктивные самки | 4,820 | 0,034 | 0,146 | 4,027 | 0,052 | |
| Между полами: | ||||||
| 3.Репродуктивные самцы и репродуктивные самки | 0,065 | 0,801 | 9,071 | 0,005 * | — | — |
| 4. Непродуктивные самцы и непродуктивные самки | 7,381 | 9 *0,025 | 0,876 | — | — | |
Мы обнаружили аналогичные эффекты размера и лечения, когда сравнивали долю драк, достигших максимального уровня интенсивности.Во всех случаях, кроме одного (самки летом, нерепродуктивный сезон, | $ P = 0,018 $ | ), не было существенной разницы в доле испытаний, достигающих максимальной интенсивности между группами малого и большого размера (репродуктивные самцы | $ P = 0,153 $ | , репродуктивные самки | $ P = 0,364 $ | , непродуктивные самцы | $ P = 0,367 $ | ). Следовательно, для трех последних курсов лечения были объединены большие и малые группы. Как и в случае продолжительности агонистических взаимодействий, значительно больше драк между репродуктивными самцами достигли максимальной интенсивности, чем между репродуктивными самками | $ (P = 0.001) $ | . Не было обнаружено существенной разницы в доле, достигающей максимальной интенсивности между непродуктивными самцами и крупными не репродуктивными самками летом | $ (P = 0,721) $ | ; тем не менее, значительно больше драк между не репродуктивными самцами достигло максимальной интенсивности, чем между маленькими не репродуктивными самками | $ (P = 0,003) $ | . Точно так же значительно больше испытаний между крупными непродуктивными самками достигло максимальной интенсивности, чем испытания между репродуктивными самками | $ (P = 0,020) $ |, в то время как не было значительной разницы в интенсивности между маленькими непродуктивными самками и репродуктивными самками | $ ( Р = 1.000) $ | . Не было существенной разницы в доле испытаний, достигающих максимальной интенсивности между самцами в репродуктивном и не репродуктивном периодах | $ (P = 0,743) $ | .
Обсуждение
Существует множество факторов, как внутренних (размер тела и чела, пол, репродуктивное состояние, предыдущий социальный опыт, мотивационное состояние, нейрохимия), так и внешних (факторы общения с окружающей средой, воспринимаемая ценность ресурсов), которые влияют на количество времени и риски для индивидов. готовы вкладывать средства в агонистические взаимодействия (Мур, 2007).Наши данные показывают, что самцы O. quinebaugensis проводили значительно больше времени в агонистическом поведении и участвовали в большей части драк с максимальной интенсивностью, чем самки в осеннем репродуктивном сезоне, как мы и предсказывали. Это может указывать на то, что самцы реагируют на половой отбор, вкладывая больше энергии в агонизм, если доминирующие самцы имеют репродуктивное преимущество.
Если доминирование действительно связано с повышенной приспособленностью самцов, можно ожидать, что в течение репродуктивного сезона, когда этот ресурс является наиболее ценным, будет инвестировано больше времени и, следовательно, больший риск.Другие исследования показали, что интенсивность и продолжительность агонистических встреч коррелируют с ценностью ресурса и его доступностью для участников (Bergman and Moore, 2003; Stocker and Huber, 2001; обзор в Moore, 2007). У раков, например, материнские самки (несущие икру или молодые особи) могут быть намного более агрессивными, чем нематериальные самки (Figler et al., 1995b), весной, когда качество и выживаемость молоди напрямую влияют на приспособленность самок. В нашем эксперименте, однако, не было значительной разницы во времени, затраченном на агонизм, или пропорции схваток, достигающих максимальной интенсивности, между репродуктивными и не репродуктивными самцами.В частности, в классах большого размера для анализа продолжительности самцы Формы II проводили столько же времени в агрессии, как и самцы Формы I. Точно так же крупные непродуктивные самки проводили больше времени в агрессии и имели более высокую долю драк с максимальной интенсивностью, чем репродуктивные самки. Хотя эта разница была значимой в отношении интенсивности борьбы, она не была значимой для времени, проведенного в агонистическом поведении, вероятно, в результате сильного эффекта размера у не репродуктивных самок.В целом, в то время как поведение репродуктивных самцов и самок согласуется с нашей гипотезой о том, что половой отбор стимулирует инвестиции в агонизм, данные по нерепродуктивным ракам — нет. Мы предлагаем два альтернативных объяснения этого наблюдения.
Во-первых, летний сезон представляет собой основной период роста как молоди, так и взрослых раков O. quinebaugensis ; такие сезонные закономерности линьки и роста обычны для раков умеренного климата (Reynolds, 2002).В этих условиях ценность таких ресурсов, как еда и жилье, может возрасти как для мужчин, так и для женщин. Монополизация пищевых ресурсов может привести к ускоренному росту и улучшению индивидуальной физической формы (Fero et al., 2007), а размер часто положительно коррелирует с результатами борьбы раков (например, Pavey and Fielder, 1996; Daws et al., 2002). ; Schroeder, Huber, 2001; Figler et al., 1995a; Klocker and Strayer, 2004; обзор в Moore, 2007) и плодовитость у самок (Rubolini et al., 2006; Reynolds, 2002), что потенциально может привести к репродуктивным преимуществам на более позднем этапе жизненного цикла для обоих полов. Укрытия также могут быть ресурсом равной ценности, который потребовал бы увеличения агонистических вложений для обоих полов, поскольку частая линька также увеличивает долю времени, в течение которого рак становится более уязвимым для хищников (Hamr, 2002). Следовательно, летом мужчины и женщины могут одинаково вкладывать средства в агонизм, потому что наиболее ценные ресурсы одинаково важны для обоих полов. Осенью относительная важность ресурсов может измениться в ответ на замедление роста и охлаждение температуры воды, а инвестиции в конкуренцию за партнеров, а не за еду или жилье отражаются в ориентированных на пол вложения в агонизм.
В качестве альтернативы, самки во время репродуктивного сезона могут извлечь большую пользу из воздержания от этих увеличенных агонистических вложений в пользу других энергетических потребностей. Для мужчин было высказано предположение, что инвестиции в агонистические взаимодействия должны конкурировать с инвестициями в другие компоненты, связанные с приспособленностью, такие как привлечение партнера или родительская забота (Qvarnström, 1997; Griffith and Sheldon, 2001), и можно было бы ожидать аналогичной торговли. снижение расхода энергии для женщин (например,г., Ветцель, 2002; Бернардо, 1996). Первичная репродуктивная деятельность самок в это время заключается в формировании яиц (Galeotti et al., 2006), что, вероятно, требует больших затрат энергии. Самки могут получить больше преимуществ в фитнесе, вкладывая больше средств в производство яиц, а не борясь за доступ к партнерам или пище, поскольку репродуктивный успех самок ограничен количеством яиц, которые она производит, а также качеством и выживаемостью ее потомства (Trivers, 1972). Было высказано предположение, что размер яйца является решающим фактором, определяющим приспособленность потомства (Bernardo, 1996), а вложение энергии в схватку с аналогами может ограничить способность самки производить большие яйца или кладку.
Поиск партнера и дискриминация также могут повлечь за собой энергетические затраты для женщин (Kokko et al., 2002). Инвестиции в агонизм в репродуктивном сезоне также могут отвлекать от времени и энергии, которые самки могут тратить на отбор и спаривание желаемых, высококачественных самцов, что в конечном итоге может повлиять на качество их потомства (Kokko et al., 2002; Berglund et al. ., 1996). У таких видов, как O. quinebaugensis , у которых самки хранят сперму и откладывают оплодотворение до нескольких месяцев после спаривания (Hamr, 2002), самки также могут получить косвенные выгоды от выделения времени и энергии на получение нескольких спариваний (Jennions and Petrie, 2000).Любые травмы, полученные в результате агрессивного поведения во время репродуктивного сезона, также могут помешать самкам спариваться с желаемым самцом или вообще успешно спариваться.
Весной, когда самка вкладывает средства в потомство в виде эмбрионов и частично независимых молодых особей, самки могут извлечь выгоду за счет повышенной агрессии, если это позволяет им оставаться в приюте, предположительно как компонент защиты потомства (Фиглер). и другие., 2001). Мур (2007) в своем обзоре соответствующей литературы обнаружил, что самки матери были более агрессивными, чем самки нематериального происхождения и репродуктивные самцы, а Фиглер и др. (2001) предположили, что это было частично связано с внутренним состоянием, поскольку даже самки матери с удаленным потомством показали более высокую защиту укрытия по сравнению с самками нематериального происхождения при сравнении предыдущих исследований. В совокупности это говорит о том, что вклад женщин в агонистические взаимодействия может происходить в рамках некоторой формы полового отбора, но в другом направлении, чем можно было бы ожидать от мужчин.
Мы также сообщаем противоречивые данные о влиянии размера на агонистическое поведение. Летом наблюдалось значительное влияние размерной группы как для самцов, так и для самок в зависимости от продолжительности: более крупные раки обоих полов проводили больше времени в агонистическом поведении, чем более мелкие раки. Также наблюдалось влияние размера на интенсивность агрессии между самками летом: у крупных самок была более высокая доля драк, достигающих максимальной интенсивности, чем у маленьких самок. Принято считать, что размер является основным фактором, который обычно предсказывает исход и агрессивное состояние при взаимодействии раков (Pavey and Fielder, 1996; Daws et al., 2002; Клокер и Страйер, 2004; Figler et al., 1995a; Шредер и Хубер, 2001; рассмотрено в Moore, 2007), и было высказано предположение, что маленькие и большие раки могут применять разные стратегии борьбы в зависимости от ценности ресурсов и риска для человека (Schroeder and Huber, 2001). Более крупные, более доминирующие люди потенциально меньше рискуют вступить в агонистическое поведение, поскольку более вероятно, что они добьются успеха, особенно в паре с меньшим противником. Однако мы не наблюдали подобных эффектов размера в брачный период, когда и большие, и маленькие особи обоих полов одинаково вкладывали средства в драки.
Хотя объяснение этому наблюдению неясно, мы отмечаем, что в нерепродуктивной коллекции животных некоторые (возможно, многие) особи обоих полов в группе меньшего размера, возможно, никогда не претерпели периода воспроизводства, в то время как особи в группах большего размера вероятность воспроизводства выше в предыдущем году. Этим можно объяснить значительную разницу, наблюдаемую в доле схваток, достигающих максимальной интенсивности, между репродуктивными самками осенью и крупными непродуктивными самками летом, в то время как сравнение репродуктивных самок осенью и мелких непродуктивных самок летом было незначительным.Материнские самки других видов раков проявляют повышенную агрессию по отношению как к репродуктивным самцам, так и к нематеринским самкам (Figler et al, 1995b), и предполагается, что это частично связано с изменением внутреннего состояния (Figler et al. ., 2001). Следовательно, возможно, что у крупных самок летом мог быть некоторый остаточный внутренний эффект, связанный с недавно рассеяной кладкой молоди, тогда как у маленьких самок меньше шансов иметь недавнюю кладку, хотя недавнее материнское состояние большинства самок в летняя группа была неизвестна.
В целом, наши данные показывают, что индивидуальные вложения в агонизм особей O. quinebaugensis различаются в зависимости от сезона, пола и размера. Тем не менее, мы специально выдвинули гипотезу о том, что взаимодействия с доминированием являются сексуально отобранным поведением, и предсказали, что репродуктивные самцы будут проявлять повышенную агрессию по сравнению как с репродуктивными самками, так и с непродуктивными раками, и наши данные не обеспечивают убедительной поддержки этой гипотезы. Вместо этого наши данные показывают, что агонистические взаимодействия у O.quinebaugensis может регулироваться сложным и динамичным набором индивидуальных затрат и выгод, и это дополнительное экспериментирование может помочь выяснить роль окружающей среды, жизненного цикла и других факторов в социальном поведении этого таксона.
Благодарности
Мы хотели бы поблагодарить Э. Райдер за ее помощь в анализе данных и следующих студентов за их помощь в сборе данных: Л. Амендола, Э. Гринуэй, К. Рикер, Р.Стэнфорд, У. Терротт, Т. Малхерн и М. Беккер. Наконец, мы хотели бы поблагодарить Инженерный корпус армии США за предоставление разрешения на специальное использование (Л. М.) для сбора раков на плотине Ист-Бримфилд в Стербридже, Массачусетс.
Список литературы
Berglund
A.
,Bisazza
A.
иPilastro
A.
1996
Вооружения и украшения: эволюционное объяснение свойств двойной полезности.
Биологический журнал Линнейского общества
58
385
—399
Бергман
DA
иМур
PA
2003
Полевые наблюдения за внутривидовым агонистическим поведением двух видов раков Orconectes rusticus и Orconectes virilis , в разных местообитаниях.
Биологический бюллетень
205
26
—35
Бергман
D.A.
иMoore
P. A.
2005
Продолжительное воздействие социальных запахов изменяет последующие социальные взаимодействия у раков Orconectes rusticus .
Поведение животных
70
311
—318
Бергман
DA
,Козловски
К.
,Макинтайр
JC
,Хубер
,Р.
,А.G.
иMoore
P. A.
2003
Временная динамика и связь эффектов победителя у раков, Orconectes rusticus .
Поведение
140
805
—825
Бернардо
J.
1996
Особый материнский эффект размера отростка, особенно размера яйца: модели, модели, качество доказательств и интерпретаций.
Американский зоолог
36
216
—236
Порода
M.D.
,Smith
S. K.
иGall
B. G.
1980
Поведенческие стратегии во время агонистических взаимодействий между самцами у тараканов.
Поведение животных
28
1063
—1069
Кандолин
U.
1999
Соревнования самцов и самцов облегчают выбор самок колюшки.
Труды Лондонского королевского общества B: Биологические науки
266
785
—789
Кокс
C.R.
иLe Boeuf
B. J.
1977
Подстрекательство самок к мужской конкуренции: механизм полового отбора.
Американский натуралист
111
317
—335
Daws
AG
,Грили
J.
,Konzen
K.
иMoore
PA
2002
Предыдущий опыт изменяет исход агрессивных взаимодействий между самцами раков, Procambarus clarkii .
Поведение и физиология в морской и пресноводной среде
35
139
—148
Fero
K.
,Simon
JS
,Jourdie
V.
иMoore
Moore
2007
Последствия социального доминирования для использования ресурсов раков.
Поведение
144
61
—82
Фиглер
M.H.
,Finkelstein
JE
,Twum
M.
иPeeke
HVS
1995a
Самцы красных болотных раков, Procambarus clarkii , сразу преобладают более мелкие представители традиционных сообществ. , смешанный сородич.
Агрессивное поведение
21
225
—236
Фиглер
M. H.
,Twum
M.
,Finkelstein
JE
иPeeke
HVS
1995b
Материнская агрессия у красных болотных раков ( Procambarus clarkii , Girard): связь между репродуктивным статусом и исходом агрессивных встреч с самцами и самками сородичи.
Поведение
132
107
—125
Фиглер
M. H.
,Blank
G. S.
иPeeke
H. V. S.
2001
Материнская территориальность как стратегия защиты потомства у красных болотных раков Procambarus clarkii , Girard.
Агрессивное поведение
27
391
—403
Фиглер
MH
,Blank
GS
иPeeke
HVS
2005
HVS
2005
Resident Shelter Procambarus clarkii Жирар и другие злоумышленники, различающиеся по полу и репродуктивному статусу.
Поведение и физиология в морской и пресноводной среде
38
237
—248
Galeotti
P.
,Pupin
F.
,Rubolini
D.
,Sacchi
,Sacchi
,Nardi
PA
иFasola
M.
2007
Влияние статуса спаривания на поведение при копуляции и расход спермы у пресноводных раков Austropotamobius italicus .
Поведенческая экология и социобиология
61
711
—718
Галеотти
P.
,Rubolini
D.
,Fea
G.
,Ghia
Nardi
P.
,Gherardi
F.
иFasola
M.
2006
Самки пресноводных раков корректируют размер яиц и кладок в зависимости от нескольких признаков самцов.
Труды Лондонского королевского общества (B)
273
1105
—1110
Griffith
S.
иSheldon
B.
2001
Фенотипическая пластичность в выражении сексуально отобранных черт: игнорируемые компоненты вариации.
Поведение животных
61
987
—993
Guiasu
R. C.
иDunham
D.W.
1998
Межформные агонистические соревнования у самцов раков, Cambarus robustus (Decapoda, Cambaridae).
Биология беспозвоночных
117
144
—154
Hagen
HA
1870
Иллюстрированный каталог Музея сравнительной зоологии Гарвардского колледжа
2
11 viii + 109 страниц
Hamr
P.
2002
Orconectes .
585
—608
Holdich
DM
Биология пресноводных раков
Blackwell Science
Oxford
Hays
WL
1994
Statistics
Harcourt
Jennions
M.
иPetrie
M.
2000
Почему самки размножаются? Обзор генетических преимуществ.
Биологические обзоры
75
21
—64
Johnsson
J. I.
1997
Индивидуальное распознавание влияет на агрессию и отношения доминирования у радужной форели, Oncorhynchus mykiss .
Этология
103
267
—282
Кирк
RE
1995
Экспериментальный план: процедуры для поведенческих наук
Brooks / Cole Publishing Company
2 Pacific Grove4,
, Pacific Grove4,
,
, Pacific Grove4,
Клокер
С.A.
иStrayer
D. L.
2004
Взаимодействия среди инвазивных раков ( Orconectes rusticus ), местных раков ( Orconectes limosus ) и местных двустворчатых моллюсков (Sphaeriidae и Unionidae).
Северо-восточный натуралист
11
167
—178
Кодрик-Браун
A.
1992
Доминирование самцов может повысить успешность спаривания у гуппи.
Поведение животных
44
165
—167
Кокко
H.
,Brooks
R.
,Jennions
M.
иMorley
J.
2002Эволюция выбора партнера и предубеждений в отношении спаривания.
Труды Лондонского королевского общества (B)
270
653
—664
Мэтьюз
Л. М.
иWarren
A. H.
2008
Новый рак из рода Orconectes Cope, 1872 г. из южной части Новой Англии (Crustacea: Decapoda: Cambariidae).
Труды Вашингтонского биологического общества
121
374
—381
Мейнард Смит
Дж.
1978
Теория оптимизации в эволюции.
Ежегодный обзор экологии и систематики
9
31
—56
Мур
P.A.
2007
Агонистическое поведение пресноводных раков — влияние внутренних и внешних факторов на агрессивные встречи и доминирование.
90
—114
Даффи
EJ
иТиль
M.
Эволюционная экология социальных и половых систем — ракообразные как модельные организмы
Harvard University Press
New York
Moore
PA
иBergman
D.A.
2005
Запах успеха и неудачи: роль внутренних и внешних химических сигналов в социальном поведении раков.
Интегративная и сравнительная биология
45
650
—657
Pavey
CR
иFielder
DR
1996
Влияние разницы в размерах на агонистическое поведение пресноводных раков Cherax cuspidatas (Decapoda: Parastacidae).
Journal of Zoology
238
445
—457
Pecor
K. W.
2006
Компромисс между преследованием и предотвращением хищничества у зрелых раков, Orconectes virilis .
Поведение и физиология в морской и пресноводной среде
39
229
—233
Qvarnstrom
A.
1997
Экспериментально увеличенный размер значка увеличивает конкуренцию мужчин и снижает заботу родителей о самцах в ошейниковой мухоловке.
Протоколы Лондонского королевского общества (B)
264
1225
—1231
Qvarnstrom
A.
иForsgren
E.
1998
Если женщины предпочитают доминирующих мужчин ?
Тенденции в экологии и эволюции
13
498
—501
Роли
MJ
иMcGuire
MT
1989
Влияние самок на доминирование самцов у 9000 обезьян в неволе7 Cercopithecus aethiops sabaeus .
Поведение животных
38
59
—67
Reichard
M.
,Bryja
J.
,Ondračková
M.
,Dávidová
,P.
иSmith
C.
2005
Половой отбор на доминирование самца снижает возможности выбора самки у европейского горчака ( Rhodeus sericeus ).
Молекулярная экология
14
1533
—1542
Рейнольдс
Дж. Д.
2002
Рост и воспроизводство.
152
—191
Holdich
DM
Биология пресноводных раков
Blackwell Science
Oxford
Rubolini
D.
,Galeotti
P.
,Ferrari
,Спайрани
М.
,Bernini
F.
иFasola
M.
2006
Распределение сперматозоидов в зависимости от мужских признаков, размера самки и поведения при совокуплении у пресноводных раков.
Поведенческая экология и социобиология
60
212
—219
Schroeder
L.
иHuber
R.
2001
Стратегии борьбы различаются размером и аллометрическим ростом раки, Orconectes rusticus .
Поведение
138
1437
—1449
Stocker
AM
иHuber
R.
2001
Стратегии борьбы с раками Orconectes rustidae состояние голода и наличие пищевых сигналов.
Этология
107
727
—736
Tilson
R.H.
иHamilton
W.J.
III1984
Социальное доминирование и особенности питания пятнистых гиен.
Поведение животных
32
715
—724
Trivers
R. L.
1972
Родительское вложение и половой отбор.
139
—179
Campbell
B.
Половой отбор и происхождение человека
Aldine
Chicago
Walker
D.
,Porter
B.A.
иAvise
J. C.
2002
Оценка генетического родства у раков Orconectes placidus , беспозвоночное с высокой плодовитостью, нуждающееся в расширенном материнском уходе.
Молекулярная экология
11
2115
—2122
Wetzel
J.
2002
Изменение формы взрослых самок раков рода Orconectes (Decapoda: Cambaridae).
American Midland Naturalist
147
326
—337
Winer
BJ
,Brown
DR
иMichels
KM
1991
Принципы статистического проектирования
McGraw-Hill
New York
Zulandt-Schneider
RA
,Huber
R.
иMoore
P.A.
2001
Распознавание индивидуума и статуса у раков, Orconectes rusticus : влияние выделения мочи на динамику схватки.
Поведение
138
137
—153
© The Crustacean Society, 2016. Опубликовано Brill NV, Leiden
Раки-мутанты, полностью самки, размножающиеся путем клонирования
Марморреба сейчас настолько распространены на Мадагаскаре, что они угрожают местным видам.Андре Карват / CC BY-SA 2.5Мраморные раки очень похожи на любых других пресноводных ракообразных. У него два когтя, десять ног и красивый сине-коричневый мраморный панцирь. Однако это шестидюймовое существо, обитающее в ручьях и озерах по всему миру, представляет гораздо более зловещие возможности, чем вы могли ожидать. Его новое научное название дает несколько подсказок: Procambarus virginalis. Все мраморные раки — самки, и они размножаются путем клонирования самих себя.
Фрэнк Лыко, биолог из Немецкого центра исследования рака, впервые услышал о мраморных раках от владельца аквариума, который в 1995 году подобрал «техасских раков» в зоомагазине.Они были поразительно большими и откладывали огромные партии яиц — сотни за один присест. Вскоре, как сообщает New York Times , любитель был осажден таким количеством раков, что раздавал их своим друзьям. И вскоре после этого marmorkrebs, как их называют по-немецки, стали появляться в зоомагазинах в Европе.
В этих раках было что-то очень странное. Все они были самками, и все они отложили сотни яиц без спаривания. Из этих яиц, в свою очередь, вылупились еще сотни самок, каждая из которых вырастает полностью способной к воспроизводству.В 2003 году ученые секвенировали их ДНК и подтвердили то, что многие владельцы уже считали так: каждый детеныш раков был клоном своей матери, и они наполняли рыбные ловушки Европы с угрожающей скоростью. «Люди начинают с одного животного, а через год у них будет пара сотен», — сказал Лико в интервью Times .
Всего 25 лет назад мраморных раков вообще не существовало. Теперь их можно найти в дикой природе миллионами в Германии, Чехии, Венгрии, Хорватии, Украине, Японии и Мадагаскаре.
Онлайн-совет для владельцев марморкребов ясен: они размножаются, как кролики. «Хотя 10 галлонов [аквариума] хватит на пару месяцев, — предупреждает один сайт, — эти 10 галлонов будут становиться все меньше и меньше по мере размножения этих животных. … Хорошее практическое правило: чем больше колония, тем больше аквариум. Начните с 40 галлонов и увеличьте объем, если можете себе это позволить ». Любители открыли эту истину на свой счет, что побудило многих выбрасывать своих дополнительных домашних животных в озера и ручьи.
В любых условиях раки прекрасно себя чувствуют.Оказывается, Marmorkrebs настолько же выносливы, насколько и плодовиты. А когда в водоеме их слишком много, они просто перемещаются. Times сообщает, что они пройдут сотни ярдов, чтобы найти новые озера или ручьи, где один экземпляр может произвести целую популяцию.
За последние пять лет Лико и его коллеги секвенировали геном мраморных раков, опубликовав свои результаты на этой неделе в журнале Nature Ecology & Evolution .Похоже, что эти раки являются результатом единственной радикальной мутации у речного рака, обитающего в реке Сатилла во Флориде и Джорджии. Около трех десятилетий назад спарились два плоских рака. При нормальном воспроизводстве каждая половая клетка имеет по одной копии каждой хромосомы. Но с одним из этих двух первых раков было что-то не так — мутация, которая оставила у него две копии в его половой клетке. Почему-то потомство оказалось выносливым и в отличном состоянии. Невероятно, но у него было по три копии каждой хромосомы и способность к бесполому размножению, не требуя участия мужчины.
Итак, захватит ли Землю армия клонов самок мраморных раков? «Это зависит от обстоятельств», — сказал Лыко « Times ». «Может быть, они просто проживут 100 000 лет. Лично для меня это займет много времени, но в процессе эволюции это будет просто отметкой на радаре ». По его словам, у их бесполого размножения есть свои преимущества и недостатки. Рост популяции стремительный, один экземпляр способен произвести сотни плодовитых потомков. Но они тоже уязвимы. При отсутствии генетических вариаций, если болезнь может уничтожить один клон, все остальные с большой вероятностью будут уничтожены.Однако сколько времени потребуется, чтобы появился такой патоген, остается загадкой. А пока во всем мире раки чувствуют себя как дома.
ADW: Cambarus bartonii: ИНФОРМАЦИЯ
Географический диапазон
Cambarus bartonii обитает в восточной и южной частях Соединенных Штатов, а также в юго-восточной части Канады. Однако чаще всего они встречаются в диапазоне от Нью-Брансуика, Канада, до северной Джорджии и в восточных частях Кентукки и Теннесси.(Зеленый, 1967)
Среда обитания
Cambarus bartonii обитают на дне ручьев, ручьев, малых рек и озер. Они строят норы, иногда называемые «дымоходами». Их норы могут быть простыми дуплами под камнем или более замысловатыми, с боковыми проходами. Дымоходы расположены вдоль кромки воды. Большая часть конструкции находится под водой, но вершина выступает и напоминает дымоход. Дымоходы различаются по размеру, самое большое отверстие составляет около восьми сантиметров.(Макман, 1960)
- озера и пруды
- реки и ручьи
Физическое описание
Cambarus bartonii — пресноводное ракообразное. При первом осмотре он выглядит «как омар». У него острая морда, а глаза на подвижных стеблях. Тонкий, но прочный экзоскелет темно-коричневого цвета, иногда со слегка красным оттенком.Точный оттенок зависит от того, как выглядит нижний субстрат среды обитания.
Из придатков наиболее заметны хелипеды. Эти придатки прикрепляются к грудной клетке и также называются первыми ходовыми ногами. Однако у этих придатков есть когти, и они используются для защиты и ловли пищи, а не для ходьбы. Cambarus bartonii также имеет четыре другие пары ходильных ног, прикрепленных к грудной клетке. Брюшные придатки Cambarus bartonii называются пловцами или плеоподами, они намного меньше ходячих ног и не подходят для плавания.Мужские пловцы модифицированы для передачи пакетов спермы самкам во время размножения и имеют лопатчатую форму. То, что похоже на веер на конце брюшка, на самом деле представляет собой множество широких плоских придатков, называемых уроподами. Все эти придатки прикреплены к брюшной стороне Cambarus bartonii , и все они двуветвистые. В целом живот большой и обычно расширенный; однако он может быть согнут под головогрудью. Первый сегмент брюшной полости обычно меньше, чем кзади от него.
Cambarus bartonii имеет открытую систему кровообращения. У него ромбовидное сердце, расположенное кпереди от брюшных сегментов по средней линии спины. Сердце окружено тонким перикардиальным мешком. Прямо перед сердцем лежат гонады. Семенники белые, а яичники оранжевые. Вдоль дорсальной средней линии головогруди лежат сердечный желудок и пилорический желудок. Сразу кзади и сбоку от желудка расположена большая пищеварительная железа.Брюшной нервный тяж лежит под внутренними органами, а мозг находится между стебельками глаз и под ними. (Рассел-Хантер, 1979; Водопич, Мур, 1999)
Репродукция
Спаривание чаще всего происходит весной, но может происходить и летом. Спаривание обычно происходит ночью, потому что шансы встретить друг друга самцом и самкой ночью в девять раз выше.
Размножение предполагает спаривание и может происходить двумя способами.Первый — это отложение спермы в семяприемник у самки. Это происходит, когда сперматозоид самца течет по бороздкам первых плеоподов и попадает в приемник самки. Сперма выходит из самца раков у основания пятой пары ходильных ног через пору. Яйца выпускаются у основания третьей пары ходильных ног. Другая форма размножения включает перенос сперматофора, и в этом случае оплодотворение является внутренним.
В любом случае оплодотворенные яйца сохраняются для созревания на плеоподах самки.Они вылупляются на плеоподах и остаются привязанными к матери вскоре после второй линьки. Говорят, что самка, несущая яйца, находится «в ягоде», потому что масса яиц похожа на ягоду. Самки чаще всего «в ягоде» в мае и июне. (Робертс, 1994; Водопич, Мур, 1999)
Поведение
Cambarus bartonii медленно ходите по дну прудов, ручьев или ручьев в поисках пищи. В случае угрозы они могут стрелять назад, чтобы быстро избежать опасности.Они совершают это быстрое движение назад, сокращая мощные мышцы живота. Cambarus bartonii используют хелипедов для защиты и поиска пищи. Хищники на C. bartonii включают различные виды птиц и черепах. Cambarus bartonii наиболее активен ночью, потому что именно тогда он питается. (Buchsbaum и др., 1987)
Привычки питания
Cambarus bartonii — хищник и падальщик.Он питается разлагающимися органическими остатками, но также ловит мелких животных. Его основными источниками пищи являются улитки, водоросли, личинки насекомых, различные виды червей и головастиков. Он находит пищу на дне источника, в котором обитает, или в почве рядом с водой. (Банистер и Кэмпбелл, 1985)
- амфибии
- яйца
- насекомые
- моллюски
- водные или морские черви
- водные ракообразные
Экономическое значение для людей: положительный результат
Раки — огромная индустрия для многих стран, особенно для Франции и США.Ежегодно потребляется от 400 000 до 800 000 кг (от 880 000 до 1 960 000 фунтов). Ежегодно в искусственных водоемах выращивают 2,6 миллиона фунтов раков. Это создает много рабочих мест для людей, занятых в рыбной отрасли.
Раки полезны и являются источником белка. Сейчас их едят в Соединенных Штатах и во многих странах мира.
Раки также могут иметь лечебные свойства. В Кении — микроскопический кровяной сосальщик.шистом вызывает инфекцию у многих людей. Шистом проникает через кожу и перемещается к мочевому пузырю и другим основным органам, вызывая серьезные повреждения. Ученые выяснили, что личинки шистома вылупляются в пресноводных улитках. Они также обнаружили, что у раков есть аппетит к этим пресноводным улиткам. Размещение большого количества раков (хотя не говорится, какие именно виды будут использоваться; у большинства видов есть аппетит к улиткам) в этих озерах может уменьшить популяцию улиток и, в свою очередь, снизить количество инфекций.
Cambarus bartonii закапывает почву вдоль ручьев, прудов и рек. Это помогает в сельском хозяйстве, потому что перекопка обогащает почву питательными веществами. На многих рисовых фермах раки являются второй культурой, потому что фермеры привозят их после уборки риса, чтобы помочь почве получить много питательных веществ. Затем фермеры собирают раков, когда снова наступает сезон риса. Это не является преимуществом для конкретного вида, потому что в мире существует множество рисовых ферм.
Раки также используются для мониторинга экологического состояния ручьев и рек. В частности, их можно проанализировать на предмет наличия в их тканях определенных загрязнителей. («Раки», 1998; Банистер и Кэмпбелл, 1985)
Экономическое значение для людей: отрицательное значение
Нет никаких известных побочных эффектов Cambarus bartonii на человека.
Статус сохранения
Известно, что этот вид не находится под угрозой исчезновения.
Авторы
Стефани Дженкинсон (автор), Юго-Западный университет, Стефани Фабрициус (редактор), Юго-Западный университет.
Глоссарий
- Неарктика
человек проживает в Неарктической биогеографической провинции, северной части Нового Света. Это включает Гренландию, канадские арктические острова и всю Северную Америку вплоть до юга до высокогорья центральной Мексики.
- двусторонняя симметрия
, имеющий такую симметрию тела, что животное можно разделить в одной плоскости на две зеркальные половины. У животных с двусторонней симметрией есть спинная и вентральная стороны, а также передний и задний концы. Синапоморфия билатериев.
- детрит
частицы органического материала от мертвых и разлагающихся организмов. Детрит — результат деятельности деструкторов (организмов, разлагающих органический материал).
- экзотермический
животных, которые должны использовать тепло, полученное из окружающей среды, и поведенческие адаптации для регулирования температуры тела
- пресная вода
в основном обитает в несоленой воде.
- гетеротермический
имеет температуру тела, которая колеблется в зависимости от температуры окружающей среды; не имеющий механизма или плохо развитый механизм регулирования внутренней температуры тела.
- гибернация
состояние, в которое некоторые животные заходят зимой, когда нормальные физиологические процессы значительно замедляются, что снижает потребность животных в энергии.Акт или условие прохождения зимы в оцепенении или состоянии покоя, обычно предполагающее отказ от гомойотермии у млекопитающих.
- итеропарное
потомков производятся более чем в одной группе (пометы, кладки и т. Д.) И в течение нескольких сезонов (или других периодов, благоприятных для воспроизводства). По определению, итеропородящие животные должны выживать в течение нескольких сезонов (или периодических изменений состояния).
- подвижный
, имеющий возможность перемещаться с одного места на другое.
- натуральный
для плавания
- родной диапазон
район, в котором животное встречается в природе, регион, в котором оно является эндемиком.
- ночной
активен ночью
- всеядное
животное, которое в основном ест всевозможные продукты, включая растения и животных
- яйцекладущие
размножение, при котором яйца выпускает самка; развитие потомства происходит вне организма матери.
- сезонное разведение
разведение привязано к определенному сезону
- малоподвижный
остается на том же участке
- половой
воспроизводство, включающее сочетание генетического вклада двух особей, мужчины и женщины
- наземный
Проживает на земле.
Список литературы
1998. Раки. Всемирная книжная энциклопедия. Лондон: World Book Inc ..
Банистер, К., А. Кэмпбелл. 1985. Омары и пресноводные раки. Стр. 236-239 в Энциклопедии водной жизни. Нью-Йорк: Equinox Ltd.
Buchsbaum, R., M. Buchsbaum, J. Pearse, V. Pearse. 1987. Животные без позвоночника. Чикаго: Издательство Чикагского университета.
Грин, Дж. 1967. Ракообразные. Стр. 172-175 в Энциклопедии жизни животных Ларусс. Нью-Йорк: Книжная компания Mc Graw-Hill.
Макман Л. 1960. Случай строительства дымохода раками * Cambarus bartonii *. Экология, 41: 383-385.
Робертс, Т. 1994. Свет, химические вещества для глаз и некоторые другие факторы как регуляторы общественной активности раков. Экологические монографии, 14: с. 376-378.
Рассел-Хантер, В.1979. Жизнь беспозвоночных. Нью-Йорк: McMillan Publishing Co Inc.
Водопич Д., Мур Р. 1999. Руководство лаборатории биологии. Берр Ридж: У. Х. Фриман и компания.
Раки (от NC WINS) | NCpedia
Камбарус бартони
Написано Сарой Пятница
Обновлено Робертом Николсом
Комиссия по ресурсам дикой природы Северной Каролины.
Классификация
Класс: Ракообразные
Отряд: Декапода «десятиногая»
Средний размер
Около 3 дюймовв длину
Еда
Питаются в основном водной растительностью, но также едят червей,
улиток, головастиков и других мелких живых или мертвых существ.
Разведение
Большинство дает молодь весной. Самка откладывает сотни яиц; вылупление около 85%. Количество выводков разное. Ученые не уверены, сколько времени требуется для вылупления яиц, но по их оценкам, месяц или больше. Когда яйца вылупляются, появляются крошечные прозрачные версии взрослых раков размером около 1/6 дюйма.долго.
Молодые
Молодь цепляется за самку через три линьки, пока не достигнет размера около 1 1/2 дюйма. Раки не проходят личиночную стадию.
Ожидаемая продолжительность жизни
Примерно 3-5 лет.
Диапазон и распространение
Некоторые раки являются универсальными и встречаются на большей части территории Северной Каролины, но большинство из них ограничено небольшими географическими районами. Раки-роющие Гринсборо, Cambarus catagius, встречаются только в районе Гринсборо, а колючие раки Северной Каролины, Orconectes carolinensis, встречаются только в бассейнах рек Тар и Нойз на прибрежной равнине.
Общая информация
Летом переверните скалу почти в любом ручье в Северной Каролине, и велика вероятность, что рак отскочит от вас назад или даже защитит скалу своими когтями, направленными на вас. Существует по крайней мере 40 различных видов этого обычного пресноводного ракообразного, которые обитают в ручьях, прудах и норах Тар Хил от гор до побережья. Многие из этих видов не встречаются больше нигде на Земле.
Описание
Раки выглядят как миниатюрные омары с передней парой сильных щипающих когтей, бронированным телом и широким хвостом.Как и у лобстеров, у раков есть 3 основные части тела: голова, грудная клетка и живот, а также твердый экзоскелет, который защищает их мягкие ткани и органы. Передняя часть тела жесткая, но задняя часть, брюшко или хвост, имеет подвижные сегменты. В области головы 2 пары антенн и крошечные глазки помогают ракам воспринимать окружающую среду. В грудной клетке у раков по 5 ног с каждой стороны (тела) и мягкие перистые жабры, которые используются для дыхания. Первая пара ног — это большие когти или клешни, которые помогают ракам ловить и удерживать добычу.Следующие 4 пары ног — это ходячие ноги; первые 2 пары — это маленькие клешни, используемые для обработки пищи. Под хвостом и после последней пары ходильных ног есть более мелкие и более тонкие придатки, похожие на ноги, называемые пловцами, которые помогают плавать, переносить яйца и удерживать молодых раков. В зависимости от вида окраска раков варьируется от малиново-красного до кобальтово-синего. Некоторые из них могут быть розовыми, коричневыми, зеленовато-черными или даже аквамариновыми.
История и статус
Наши биологи ежегодно открывают и описывают новые виды раков, а также документируют распространение экзотических раков в штате.По мере завершения новых исследований биологи продолжают документировать расширение ареала, а также сокращение некоторых раков Северной Каролины.
Среда обитания и привычки
Раки обитают в ручьях, прудах, озерах и болотах по всей Северной Каролине. Обитатели ручьев предпочитают быстро движущиеся и насыщенные кислородом реки и ручьи гор и предгорий. В более медленных ручьях, например на Прибрежной равнине, раки прячутся под камнями и бревнами для защиты.Раков также можно встретить в прудах, озерах и в стоячей воде в придорожных канавах. Некоторые раки зарываются в землю и могут рыть разветвляющиеся туннели глубиной до 15 футов в илистых заболоченных местах. Большинство нор в среднем составляют 2–3 фута в глубину, и их можно найти на приличном расстоянии от стоячей воды. Раки-роющие, обитающие в основном на Прибрежной равнине и в восточной части Северной Каролины, во время копания часто выбрасывают глиняные «дымоходы».
Раки становятся наиболее активными с наступлением темноты или на рассвете и выходят из своих укрытий, чтобы покормиться.Эти стойкие ракообразные питаются в основном водной растительностью, но они также могут есть мелких, живых или мертвых животных, таких как черви, улитки, головастики и рыба. Раки часто становятся добычей других животных, таких как лягушки, черепахи, окуни, еноты, болотные птицы и ястребы. Но раки — бойцы. Лучшая защита — мощный хвост, который отбрасывает раков назад. Если они теряют коготь или ногу в бою, на их месте вырастает новый.
Большинство видов раков в Северной Каролине дают потомство ранней весной, когда много еды и дождя.Некоторые виды размножаются зимой. Для спаривания самец рака хватает самку и переворачивает ее так, чтобы брюшная сторона находилась вместе. Затем самец передает сперму самке через свою первую пару плавников, называемых гоноподом. При оплодотворении сотни крошечных яиц прикрепляются к плавникам самки с липкими выделениями, называемыми яицами. Когда к самке раков прикрепляются оплодотворенные яйца, говорят, что она в ягоде. Более крупные самки обычно откладывают больше яиц.
В отличие от многих других ракообразных, раки не проходят личиночную стадию, а линяют около 6 раз в первые 7-8 месяцев жизни.Линька или выпадение панциря необходимо, потому что раки перерастают свою броню. Они продолжают линять более медленными темпами всю оставшуюся жизнь. Когда они линяют, старый скелет отпадает, и раки с мягким панцирем быстро растут, так как поглощают большое количество воды. Незащищенные, они прячутся после линьки и менее агрессивны, пока через несколько дней не затвердеет новый скелет. Раки прекрасно себя чувствуют в незагрязненной воде, и их количество является хорошим признаком хорошей экологии.
Взаимодействие с людьми
Раки ценятся не только с экологической точки зрения, но и с экономической точки зрения.Более крупные раки, такие как красные болотные раки, считаются деликатесом каджунской кухни. Раки также используются в качестве приманки для рыбной ловли, и это привело к некоторым случайным проникновениям неместных раков в новые районы. Неместные раки вредят нашей экосистеме, конкурируя с местными видами тархилов и уничтожая водные растения, которые служат пищей и убежищем для местных животных. ПОЖАЛУЙСТА, НЕ выпускайте раков в водные пути Северной Каролины !!! Попав сюда, их невозможно искоренить! Вот совет для тех, кому не посчастливилось попасть не на тот конец раков.У них есть острый укус, но он укусил их только в качестве последней попытки самообороны. Если им дадут шанс сбежать, они это сделают. Так что, если вы когда-нибудь обнаружите, что к вам прицепились раки, просто
воткните пораженную часть анатомии обратно в воду, где раки будут чувствовать себя в большей безопасности и уплывут. Этот метод также работает с черепахами.
Примечание: «раки» относятся к одному виду или отдельному экземпляру, тогда как «раки» относятся ко многим видам.
Судьба золотистых желез и нерестовое поведение у неместных раков: сравнение гонохорических речных раков и партеногенетических мраморных раков.
Abstract
В период до нереста у самок пресноводных раков развиваются золотистые железы на нижней стороне плеона.Эти железы производят слизь для временной структуры, похожей на палатку, в которой яйца оплодотворяются и прикрепляются к плеоподам. Долгосрочное наблюдение за самками рака Procambarus fallax , размножающихся двуполым путем, содержащихся в неволе, показало, что голые железы развиваются в конце зимы и в конце лета каждого года независимо от присутствия самцов. Однако, в отличие от спарившихся самок, самки без спаривания не образуют ни оплодотворяющей палатки, ни нереста. Их голые железы сохранялись необычно долго и исчезли только во время следующей линьки.Судя по всему, женщины используют информацию о наличии сперматозоидов либо для воспроизводства, либо для сохранения ресурсов. Мраморный рак, Procambarus virginalis , партеногенетический потомок слепого рака, развил голые железы примерно в одни и те же периоды года, но нерестился, несмотря на отсутствие спаривания. Эти находки указывают на то, что на пути от гонохоризма к партеногенезу регуляция активности голых желез и нереста была отделена от спаривания. Таким образом, пара видов Procambarus fallax / Procambarus virginalis кажется особенно подходящей для исследования физиологических, молекулярных и генетических механизмов, лежащих в основе нереста пресноводных раков.
Введение
За несколько недель до нереста у пресноводных раков на нижней стороне плеона развиваются выступающие белесые блестящие железы (Reynolds 2002). Эти железы сначала выглядят как слабые кремово-беловатые пятна и становятся все более и более отчетливыми, что является хорошим индикатором предстоящего нереста (Jussila et al. 1996). Незадолго до откладывания яиц голые железы выделяют полупрозрачную слизь, которая образует тентовую структуру на нижней стороне грудной клетки и плеона (Andrews 1904; Vogt 2016).В этой студенистой массе сперматозоиды мобилизуются из сперматофоров, которые прикреплены или хранятся внутри, в зависимости от семейства раков (Reynolds 2002; Vogt 2002; Niksirat et al. 2013). Палатка для оплодотворения дополнительно обеспечивает прикрепление оплодотворенных яиц к плеоподам, где они высиживаются в течение нескольких недель до вылупления первой молоди и далее (Vogt and Tolley 2004). Глэровые железы и палатка оплодотворения являются уникальными особенностями пресноводных раков и встречаются во всех трех семействах: Astacidae, Cambaridae и Parastacidae (Andrews 1906; Mason 1970; Thomas and Crawley 1975; Jussila et al.1996; Фогт 2016). Более простые цементные железы, которые только способствуют прикреплению яиц к плеоподам, встречаются и у других Decapoda (Adiyodi and Anilkumar 1988).
Во время конференций по ракообразным и студенческих курсов я часто сталкивался с вопросом, что происходит с желчными железами и зрелыми ооцитами, если самки раков не спариваются. К сожалению, нет опубликованных данных о судьбе желчных желез и очень мало данных о нересте у несвязанных самок (Woodlock, Reynolds 1988; Reynolds et al.1992; Холдич и др. 1995; Tropea et al. 2010; Buřič et al. 2011). Одна из возможностей состоит в том, что несвязанные самки нерестятся как спарившиеся самки, но неоплодотворенные яйца разлагаются. В качестве альтернативы слизь и зрелые ооциты могут реабсорбироваться, чтобы избежать растраты ресурсов. В настоящем сообщении я сообщаю о судьбе головых желез и нерестовом поведении у выращенных в лаборатории спарившихся и не спарившихся речных раков Procambarus fallax и их партеногенетически воспроизводящегося родственника, мраморного рака Procambarus virginalis .У мраморных раков нет самцов, и они произошли от речных раков в результате аутотриплоидизации и сопутствующего перехода от гонохоризма к партеногенезу (Scholtz et al. 2003; Martin et al. 2010; 2016; Vogt et al. 2015). Недавно они были признаны отдельным видом из-за репродуктивной и географической изоляции от Procambarus fallax , уникальных генетических и эпигенетических признаков, а также превосходного роста и плодовитости (Vogt et al. 2015).
Материалы и методы
Самки речного рака Procambarus fallax (Hagen 1870) (Decapoda: Cambaridae) были отделены от своих сородичей-самцов на пятой стадии молоди, первой стадии жизни, которая позволяет определить пол по внешнему признаку.В последующие пять месяцев их держали вместе в женской группе, а затем по отдельности. Три из этих самок остались без спаривания, а еще три самки были повязаны с самцами в течение сезонов размножения весной 2015, осенью 2015 и весной 2016. Спаривание производилось путем спаривания отдельных самок с гладкими железами и отдельных самцов с крючками на седалищных костях 3-й и 3-й седалищных желез. 4-е переоподы (показатели половой зрелости) в контейнере 30 × 25 × 20 см. Копуляция считалась успешной, когда самцы переворачивали самок на спину и вставляли их копулятивные органы в приемник спермы (annulus ventralis) самки не менее чем на 30 минут.Все спарившиеся и не спарившиеся самки содержались индивидуально в пластиковых контейнерах 30 × 25 × 20 см, снабженных гравием и укрытием. В качестве источника воды использовалась водопроводная вода, которую меняли раз в неделю. Температура воды колебалась от ~ 15 ° C зимой до ~ 25 ° C летом, и фотопериод был естественным режимом окружающей среды Гейдельберга (Германия). Раков кормили ежедневно ad libitum гранулами TetraWafer Mix. Период наблюдений длился с 1 июля 2014 г. по 31 мая 2016 г.Я дал время в неделях только потому, что в начале созревания слепой железы трудно решить, началось ли уже развитие или нет. Данные по Procambarus fallax были сопоставлены с моими записями о наличии блестящих желез у партеногенетических мраморных раков Procambarus virginalis , которые я собирал за последние 13 лет. Мраморных раков выращивали в тех же условиях, что и речных раков, индивидуально или коллективно.
Результаты
У самок обоих видов раков в период размножения развиваются голые железы. Эти железы были расположены в последней грудной части грудины, грудины и плевре плеона, уропод и плеопод (рис. 1А, В). Головые железы развивались автономно, в отсутствие самцов.
Рис. 1.Глеровские железы и приемник для сперматозоидов у раков. (A) Зрелые железы голени на нижней стороне плеона партеногенетической самки Procambarus virginalis .Беловатые железы расположены в грудины (стрелка), плевре (наконечник стрелки), уроподах (u) и плеоподах (p). Пруток: 5 мм. (B) Плеон самки Procambarus fallax без спаривания с обнаженными железами через 12 недель после их первого появления. Пруток: 2 мм. (C) Сканирующая электронная микрофотография брюшного кольца самки Procambarus virginalis . Бар: 200 мкм.
В трех проанализированных периодах воспроизводства нерестовые самки Procambarus fallax не спаривались, хотя у них регулярно развивались голые железы (Таблица 1).Напротив, спарившиеся самки обычно откладывали яйца. В последнем случае блестящие железы были видны в течение трех-пяти недель. Их опорожняли незадолго до кладки яиц, чтобы сформировать палатку для оплодотворения. У несвязанных самок золотистые железы сохранялись до 12 недель (таблица 1) и исчезли только во время следующего шелушения.
Таблица 1.Судьба золотистых желез и нерестовое поведение у не спарившихся и не спарившихся Procambarus fallax самок.
Мраморных раков, выращиваемых в лаборатории, в основном воспроизводятся два раза в год, как и речных раков (Vogt 2015).Некоторые особи размножаются только один-три раза в год. За 3-6 недель до нереста у них были обнаружены железы с увеличивающейся интенсивностью окрашивания. В отличие от несвязанных самок речного рака, партеногенетические самки мраморных раков производили палатки для оплодотворения и нерестились. Только в одном из 103 задокументированных случаев самка с голыми железами пропустила нерест и вместо этого выполнила линьку. Золотистые железы этого экземпляра исчезли во время линьки, как и у несвязанных раков.
Обсуждение
Сравнение не спарившихся самок гладкоплодных раков и партеногенетических мраморных раков выявило интересные различия в отношении нереста и судьбы гладких желез.Спаривающиеся самки и самки мраморных раков обычно нерестятся дважды в год (Vogt 2015). Эти нереста связаны с опорожнением желёз. У слабых раков оба события зависят от спаривания. Несвязанные Procambarus fallax самок не порождают и не опорожняют свои голые железы. Вместо этого блестящие железы сохраняются еще много недель, вероятно, в ожидании более позднего шанса спаривания. Это контрастирует с мраморными раками, у которых развиваются голые железы и нерестятся в отсутствие самцов.Очевидно, что информационная система, позволяющая различать условия спаривания и отсутствия спаривания, была потеряна или изменена во время перехода от гонохоризма к партеногенезу в паре видов Procambarus fallax / Procambarus virginalis .
Полное подавление нереста наблюдалось также у самок астцид Austropotamobius pallipes , Astacus leptodactylus и Pacifastacus leniusculus по данным Holdich et al.(1995). Напротив, Вудлок и Рейнольдс (1988) наблюдали нормальный нерест, но распад прикрепленных яиц у несвязанных самок Austropotamobius pallipes , которые содержались в парах самок. У самок выращенных в лабораторных условиях Cherax destructor (Parastacidae), не имеющих спаривания, нерестовое поведение было весьма разнообразным: около 20% нерестились при выращивании при 30 ° C, но не появлялись при 27 ° C (Tropea et al. 2010). Нерест не спарившихся самок произошел и у факультативно партеногенетических самок камбарид Orconectes limosus (Buřič et al.2011, 2013). Из 30 не спаривавшихся партеногенетических самок 28 нерестились, и только одна самка не отнерестилась (оставшаяся особь погибла) (Buřič et al. 2011), как и у мраморных раков. Причины различий в нерестовом поведении самок, не имеющих спаривания, между указанными размножающимися половым путем видами и внутри них неизвестны, но могут зависеть от видовых различий, условий окружающей среды, предэкспериментального опыта и статуса питания особей. Информация о судьбе желёзок отсутствует во всех цитированных выше сообщениях.Подавление секреции и нереста гололеда можно интерпретировать как энергосберегающую стратегию.
Созревание ооцитов не зависит от присутствия самцов, как показали все вышеупомянутые исследования. То же самое касается созревания желёзок, которые развиваются за несколько недель до нереста. Оба процесса, по-видимому, тесно связаны и вызываются факторами окружающей среды, в основном температурой и световым периодом (Reynolds 2002; Vogt 2015), которые влияют на гормональную систему. Созревание яичников ингибируется гормоном, ингибирующим вителлогенез (VIH) из системы X-орган-синусовая железа, и стимулируется гормоном, стимулирующим яичники (OSH), из ганглиев грудных нервов (Reynolds 2002; Vogt 2002).Созревание блестящих желез, вероятно, регулируется теми же гормонами.
Спаривание может быть одним из спусковых механизмов для нереста, как предполагает Рейнольдс (2002). Это предположение подтверждается экспериментами Holdich et al. (1995), которые спарили самок с самцами другого вида. Такие самки обычно нерестились, но икринки не развивались. Однако в естественных условиях между спариванием и нерестом часто бывает много дней или даже недель, что позволяет предположить, что нерест также может зависеть от воспоминаний о спаривании самкой или ощущения присутствия спермы.У астацидных и парастакидных раков сперматофоры прикреплены снаружи к грудным пластинам грудины и хвостовому вееру (Reynolds 2002; Vogt 2002; Niksirat et al.2013), и, таким образом, самки могут легко ощущать присутствие сперматозоидов своими переоподами. У Cambaridae сперма хранится внутри вентрального кольца (рис. 1C) (Reynolds 2002; Vogt et al. 2004), и в этом случае механорецепторы могут дать информацию о состоянии ее наполнения. №
Неизвестно, могут ли раки добровольно опорожнять железы и яичники за счет сокращения мускулатуры желез и яичников.Более вероятно, что оба процесса являются непроизвольными и регулируются гормонами, такими как простагландин F 2α , который, как было показано, резко усиливается на заключительной стадии оогенеза у Procambarus paeninsulanus (Spaziani et al. 1995). Тесты in vitro показали, что этот гормон вызывает сокращение ткани яичников в зависимости от дозы.
Настоящее исследование является первым отчетом о судьбе железистой оболочки без спаривания раков, выращенных в контролируемых условиях.