Структурная схема и принцип работы микросхемы управления импульсными источниками питания
Основной недостаток рассмотренных выше микросхем линейных стабилизаторов — это большая мощность, рассеиваемая на проходном транзисторе, что ограничивает область их применения. Основное достоинство импульсных источников питания заключается в их способности обеспечить большую мощность в нагрузке, имея при этом высокий коэффициент полезного действия (КПД) и малые габариты. В отличие от линейных стабилизаторов, в импульсных источниках питания проходной транзистор переключается с большой частотой (от 20 кГц до 5,0 МГц). Ключевой транзистор коммутирует индуктивный элемент (обмотку трансформатора либо дроссель), в котором накапливается энергия, пропорциональная времени открытого состояния транзистора. После закрытия ключевого транзистора энергия, накопленная в индуктивном элементе, передается в нагрузку. Благодаря тому, что на проходном транзисторе не происходит падения напряжения Um — t/bix, импульсные ИМС рассеивают гораздо меньшую мощность по сравнению с линейными стабилизаторами.
Большая частота переключения позволяет использовать индуктивные элементы малых значений и, соответственно, малых размеров. Кроме того, пульсирующее напряжение большой частоты можно «сгладить» емкостью небольшого значения. В качестве ключа обычно используется мощный, быстродействующий MOSFET, который по своим характеристикам предпочтительнее биполярного транзистора. Изменением времени включения MOSFET производится регулирование количества энергии, передаваемой во вторичную цепь так, что выходное напряжение источника питания остается независимым от изменения нагрузки.
Поскольку сигнал управления мощным транзистором, выдаваемый с микросхемы, представляет собой последовательность прямоугольных импульсов с шириной, пропорциональной управляющему сигналу, то такие микросхемы называют еще ШИМ-контроллерами [19], или ИМС управления импульсными источниками питания [22]. Типовая структурная схема, поясняющая работу импульсного источника питания, приведена на рис.
Блок выпрямителя сглаживает пульсации сетевого напряжения. Микросхема контроллера импульсным источником питания, как правило, включает в себя источник питания с блоками опорного напряжения (£/оп), усилитель сигнала ошибки (УСО), ШИМ-компаратор, генератор пилообразных напряжений, блок логики, формирователь управляющих напряжений, блок защиты от повышенной температуры кристалла t °С, повышенного выходного тока и тока короткого замыкания, повышенного и пониженного напряжения питания сети.
Источник питания ИМС содержит в своем составе источник опорного температуронезависимого напряжения. Служит источник питания для формирования напряжения питания всех блоков ИМС и для выработки опорного напряжения.
Рис. 3.23. Схема импульсного источника питания с использованием ШИМ-контроллера: УСО — усилитель сигнала ошибки; ГПН генератор пилообразного напряжения; ЛСН — линейный стабилизатор напряжения
Опорное термостабилизированное напряжение подается на усилитель сигнала ошибки, на второй вход которого подается выпрямленный сигнал со вторичной обмотки импульсного трансформатора. Усиленная разница сигналов поступает на ШИМ-компаратор. Это напряжение сравнивается с напряжением, вырабатываемым генератором пилообразных напряжений (ГПН). На выходе ШИМ-компаратора появляется прямоугольный сигнал в случае, если пилообразное напряжение превышает напряжение сигнала ошибки 1/ош. Таким образом, чем выше напряжение сигнала t/ouj, тем меньшая длительность импульса на выходе ШИМ- компаратора. Так происходит широтно-импульсная модуляция. Этот сигнал проходит блок логики, усиливается и подается на затвор мощного MOSFET-транзистора. MOSFET может быть отдельным полупроводниковым прибором или может входить в состав ИМС.
Усиленная разница сигналов поступает на ШИМ-компаратор. Это напряжение сравнивается с напряжением, вырабатываемым генератором пилообразных напряжений (ГПН). На выходе ШИМ-компаратора появляется прямоугольный сигнал в случае, если пилообразное напряжение превышает напряжение сигнала ошибки 1/ош. Таким образом, чем выше напряжение сигнала t/ouj, тем меньшая длительность импульса на выходе ШИМ- компаратора. Так происходит широтно-импульсная модуляция. Этот сигнал проходит блок логики, усиливается и подается на затвор мощного MOSFET-транзистора. MOSFET может быть отдельным полупроводниковым прибором или может входить в состав ИМС.
На блок логики поступает сигнал с блока защит. Как правило, в ИМС управления импульсными источниками питания имеются схемы защиты от повышенной температуры кристалла, от повышенного тока нагрузки и короткого замыкания в нагрузке, от скачков сетевого напряжения по верхней границе OVP и по нижней границе.
Источник: Белоус А.И., Ефименко С. А., Турцевич А.С., Полупроводниковая силовая электроника, Москва: Техносфера, 2013. – 216 с. + 12 с. цв. вкл.
А., Турцевич А.С., Полупроводниковая силовая электроника, Москва: Техносфера, 2013. – 216 с. + 12 с. цв. вкл.
Как устроены интегральные схемы
- Дата
- Категория: it
Компьютеры строятся на основе двух типов интегральных схем: логической и ЗУ (запоминающее устройство). Логические микросхемы используются в арифметическом логическом модуле (АЛМ), где производятся вычисления, в то время как кристаллы ЗУ хранят данные и программы. Существует множество разновидностей логических микросхем, простых и сложных; микропроцессорная схема (нижняя правая иллюстрация на стр. 23) выполняет роль центральной нервной системы ПК и является ярким примером сложной логической схемы. Иногда функции логических схем и ЗУ комбинируются в одной схеме.
Микропроцессор служит центральным процессорным устройством (ЦПУ) компьютера, включающим контроллер и оперативную логическую схему. Другие, примыкающие к процессору схемы включают: генератор синхроимпульсов, который производит сигналы, обеспечивающие пошаговую деятельность компьютера; контроллер ввода/вывода, который координирует ввод и вывод данных; различные сопроцессоры — процессоры, специализированные для одного вида задач и выполняющие их с огромной скоростью. Дополнительные контроллерные схемы оперируют со связующими схемами, магнитными дисками и графическими терминалами.
Дополнительные контроллерные схемы оперируют со связующими схемами, магнитными дисками и графическими терминалами.
Устройство интегральной схемы
величенная в 2500 раз структура МОП — металл-оксид-полупроводник для отрицательного канала -является распространенным типом ИС транзистора. Обычно этот переключатель закрыт; ток (голубая стрелка) не может пройти от источника к стоку. Но напряжение (красная стрелка), примыкающее к логической схеме, притягивает электроны (точечки), образуя канал, который пропускает электрический ток.
Схемы ИС в корпусах
Прежде чем ИС будет смонтирована на печатной плате, она должна быть заключена в защитный футляр, или корпус, и снабжена внешними связующими штырями, или выводами. На иллюстрации справа представлено несколько разновидностей корпусов, получивших свое название по форме и организации выводов корпуса. DIP — переключатель, или двухрядный корпус, имеет два ряда выводов. PLP — это уплощенный корпус, с выводами по двум сторонам. LCC — керамический кристаллодержатель без выводов. ZIP — плоский корпус со штырьковыми выводами, расположенными зигзагообразно. QFP — это плоский корпус с четырьмя рядами выводов по бокам. SIP — корпус с однорядным расположением выводов.
Монолитный микропроцессор
Схема на изображении внизу включает микропроцессор, а также схемы ПЗУ, ОЗУ и контроллеры (регуляторы ввода/вывода). Эти компьютерные схемы широко применяются для управления машинным оборудованием и многими бытовыми приборами.
Ряды схем.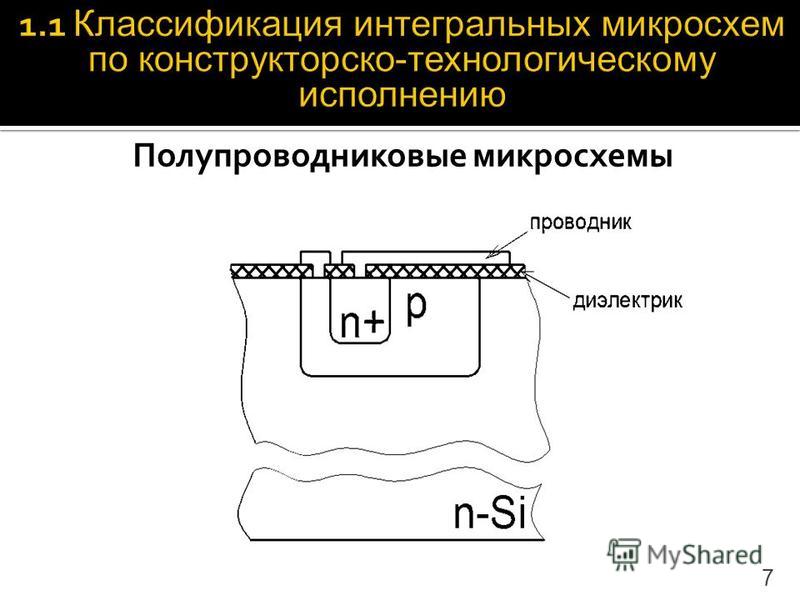 Печатная плата на илл. слева, объединительная плата ПК, содержит несколько видов ИС, включая микропроцессор, контроллерные схемы и ЗУ.
Печатная плата на илл. слева, объединительная плата ПК, содержит несколько видов ИС, включая микропроцессор, контроллерные схемы и ЗУ.
Микросхемы ШИМ-контроллера UC3844, UC3845, UC2844, UC2845
Микросхемы ШИМ-контроллера UC3844, UC3845, UC2844, UC2845 являются самыми распространенными в импульсных блоках питания бытовой и компьютерной техники, используется для управления полевым ключевым транзистором в схемах импульсных блоков питания. Они специально разработаны для DC− DC преобразователей — преобразование постоянного напряжения одной величины в постоянное напряжение другой величины.
Принцип работы микросхем UC3844, UC3845, UC2844, UC2845
Принцип работы микросхемы UC3844: При напряжении питания в норме, на выводе 8 появляется напряжение +5В, которое запускает генератор OSC , генератор в какой-то момент выдает короткий положительный импульс на вход RS, S триггера, переключая его, после этого на выходе появляется нуль. При спаде импульса OSC, напряжение, на прямых входах цифрового элемента станет равным нулю.
Рис. 2. Структурная схема микросхем UC3844, UC3845, UC2844, UC2845, в скобках указаны номера выводов микросхем в 14ти выводных корпусах (с суффиксом D, см. цоколевку выше).
При этом, на инвертирующем выходе образуется логическая 1, эта единица откроет верхний транзистор, и ток от плюс источника, коллектор, эмиттер потечет в нагрузку подключенной к выходу (6 вывод). Импульс на выходе будет открытым и длится до тех пор, пока на вывод 3 не поступит закрывающее напряжение выше +1 Вольт. При подачи напряжения на 3 вывод (выше +1 Вольт), и на прямой вход операционного усилителя, на выходе появится логическая 1, и переключит RS триггер в момент подачи (лог. 1) на вход R. В результате на выходе RS триггера появится логическая единица, при подачи еденицы на один, из прямых входов логического элемента, на его прямом выходе образуется логическая единица (на инверсном выводе в этот момент образуется логический 0, запирающий верхний транзистор), в результате открывает нижний транзистор и через коллектор-эмиттер замыкает выход (вывод 6 микросхемы) на «землю».
Типовые схемы включения микросхем UC3844, UC3845, UC2844, UC2845
На схемах, в скобках указаны номера выводов микросхем в 14ти выводных корпусах (с суффиксом D, см. цоколевку выше).
Пример реализации импульсного блока питания на на базе ШИМ-контроллера UC3844
Принципиальная схема импульсного блока питания на базе ШИМ-контролера UC3844 и силовом ключе на полевом транзисторе STP3NA90F.
Микросхемы с наименованием UC3844 кроме UNITRODE выпускают фирмы ST и TEXAS INSTRUMENTS, аналогами этой микросхемы являются: DBL3844 фирмы DAEWOO, SG3844 фирмы MICROSEMI/LINFINITY, KIA3844 фирмы КЕС, GL3844 фирмы LG, а также микросхемы других фирм с различными литерами (AS, МС, IP и др.) и цифровым индексом 384Х.
Логический элемент 2И-НЕ ТТЛ
(микросхема К155ЛА3)
Соответствие выводов микросхемы К155ЛА3 светодиодам испытательного стенда
показано на рисунке 2. 5, а условное обозначение микросхемы — на рисунке 2.6.
5, а условное обозначение микросхемы — на рисунке 2.6.
Задания:
1. Вставить печатную плату с микросхемой К155ЛА3 в разъем испытательного стенда.
2. Подключить испытательный стенд к источнику постоянного напряжения 5 В с соблюдением полярности (провод в изоляции красного цвета подсоединяют к выводу «+» источника).
3. Проверить работоспособность логических элементов микросхемы К155ЛА3, записав в тетради напряжение логического нуля и логической единицы для каждого элемента. Составить таблицу истинности элемента.
4. Вынуть проверенную микросхему из панельки и проверить работоспособность еще одной микросхемы К155ЛА3. Вставляя проверяемую микросхему в панельку, необходимо убедиться в соответствии первого вывода микросхемы (определяется по ключу на микросхеме) первому выводу панельки (определяется по ключу на панельке).
5.
Объяснить
принцип работы логического элемента 2И-НЕ микросхемы К155ЛА3 по принципиальной
схеме, приведенной на рисунке 2. 7. Указать назначение всех элементов схемы,
обратив особое внимание на резисторы R4, R5 и транзистор VT5. Нарисовать схемы подключения
нагрузки для случаев логической единицы и логического нуля на выходе элемента.
Для обоих случаев показать цепи, по которым протекает ток нагрузки.
7. Указать назначение всех элементов схемы,
обратив особое внимание на резисторы R4, R5 и транзистор VT5. Нарисовать схемы подключения
нагрузки для случаев логической единицы и логического нуля на выходе элемента.
Для обоих случаев показать цепи, по которым протекает ток нагрузки.
6. Вычертить в тетради контактно-релейные схемы логического элемента 2И-НЕ для режимов положительной и отрицательной логики. Для контактно-релейных схем в режиме положительной логики логической единице соответствует замкнутый ключ, горящая лампочка также соответствует логической единице. В режиме отрицательной логики логической единице соответствует разомкнутый ключ, не горящая лампочка указывает на состояние логической единицы.
Принцип работы микросхемы 4046 (К564ГГ1) для устройств с удержанием резонанса
Принцип работы микросхемы 4046 (К564ГГ1) для устройств с удержанием резонанса
Микросхема К561ГГ1 представляет собой генератор с ФАПЧ. Применяется для частотной и фазовой модуляции, демодуляции ЧМ-сигналов, тактовой синхронизации и синтеза частоты, а также для умножения частоты.
Применяется для частотной и фазовой модуляции, демодуляции ЧМ-сигналов, тактовой синхронизации и синтеза частоты, а также для умножения частоты.
При создании силового электронного устройства с удержанием резонанса в LC-контуре, для синхронизации полученных колебаний с управляющими импульсами, идущими от драйвера, проектируют схему резонансного контроллера.
Задача данного контроллера — удержать резонансные колебания в LC-контуре посредством его возбуждения в такт собственным колебаниям. Чтобы этого добиться, контроллеру необходимо по цепи обратной связи получать от контура сигнал, содержащий данные о текущей частоте и фазе свободных колебаний в нем, после чего, с опорой на эти данные, поддерживать работу драйверного каскада синхронно с этими частотой и фазой, тогда и резонанс в контуре будет автоматически сохраняться.
Для построения такого контроллера подходит микросхема CD4046 или ее отечественный аналог К564ГГ1. Давайте рассмотрим устройство этой микросхемы, назначение ее выводов и схему подключения навесных компонентов, чтобы при необходимости понимать, с чем имеешь дело.
Данная микросхема позволяет легко организовать цепь ФАПЧ — фазовой автоподстройки частоты. Для построения ФАПЧ здесь используются три необходимых блока, расположенные внутри микросхемы: ГУН — генератор, управляемый напряжением, ФК — фазовый компаратор и ФНЧ — фильтр низкой частоты.
Встроенный в микросхему, ГУН выдает последовательность прямоугольных импульсов с заполнением 50%, то есть чистый меандр, начальная частота которого зависит от параметров двух RC-цепей: R1C1 и R2C2, присоединенных к нему снаружи микросхемы, а амплитуда в данном случае приближена к напряжению питания микросхемы U+.
Принцип работы ФАПЧ
Внешний входной сигнал fвх подается на микросхему, на самом деле — на один из входов фазового компаратора ФК (ФК1 или ФК2 — выбирает разработчик) внутри нее. На второй вход ФК параллельно подается меандр, вырабатываемый ГУНом. В результате, на выходе ФК получается прямоугольный сигнал, длительность импульсов в котором зависит от разницы между импульсами с ГУНа и внешними импульсами в каждый момент времени.
По сути, длительность выходных импульсов с ФК пропорциональна разности фаз двух сравниваемых сигналов. Дело в том, что в роли ФК часто используется логический элемент «исключающее ИЛИ», это значит, что на выходе ФК высокий уровень напряжения будет лишь в том случае, если между сигналами есть разница, а если разницы нет то на выходе с ФК будет низкий уровень напряжения или состояние бездействия.
С выхода ФК сигнал подается на фильтр низкой частоты, представляющий собой простую RC-цепь, на конденсаторе которой получается пульсирующее напряжение рассогласования, причем уровень пульсаций оказывается пропорционален разнице двух сигналов (от внутреннего ГУН и подаваемого на микросхему извне), по сути — разности их фаз.
Полученное на конденсаторе ФНЧ, напряжение рассогласования тут же подается обратно на вход ГУН, и в зависимости от его средней величины частота ГУН будет автоматически перестраиваться так, чтобы частота меандра на его выходе fвых приближалась бы к частоте внешнего сигнала, приходящего на микросхему извне. По достижении такой ситуации, среднее напряжение на конденсаторе фильтра низкой частоты будет наименьшим — это и есть признак наступления максимального сближения двух сигналов по частоте и фазе. Когда сигнал таким образом захвачен, он продолжит удерживаться петлей ФАПЧ.
По достижении такой ситуации, среднее напряжение на конденсаторе фильтра низкой частоты будет наименьшим — это и есть признак наступления максимального сближения двух сигналов по частоте и фазе. Когда сигнал таким образом захвачен, он продолжит удерживаться петлей ФАПЧ.
Пределы перестройки ГУН
Как вы уже поняли, частота ГУН способна перестраиваться в пределах определенного диапазона автоподстройки. Данный диапазон задается внешними компонентами микросхемы. А скорость реакции системы ФАПЧ определяется постоянной времени НЧФ (величинами C2 и R3). По этой причине следует строго подходить к выбору навесных компонентов микросхемы.
Напряжение питания микросхемы, конденсатор C1, а также резисторы R1 и R2 определяют диапазон автоподстройки частоты ГУН внутри микросхемы. Резистор R2 смещает минимальную частоту fmin ГУН выше от нуля. А соотношение между номиналами резисторов R1 и R2 определяет соотношение между максимальной и минимальной частотами — fmax/fmin, перестраиваемого выходного сигнала с ГУН.
Входы и выходы микросхемы
Вывод 4 — сигнальный выход ГУН, на нем в рабочем режиме меандр. Данный выход можно использовать для подачи сигнала к другим блокам проектируемого устройства.
Вывод 5 отвечает за включение и выключение ГУН. При подаче на данный вывод напряжения высокого уровня, микросхема отключится. При подаче напряжения низкого уровня (при подключении вывода 5 к общему проводу) — микросхема будет работать в штатном режиме.
Выводы 6 и 7. К ним подключается конденсатор C1 — это частотозадающий конденсатор ГУНа.
Вывод 8 — общий провод питания микросхемы.
Резистор R1 – между выводом 11 и общим проводом. Резистор R2 – между выводом 12 и общим проводом. Это частотозадающие резисторы. Резистор R3 фильтра НЧ – к выводу 9 и выводу 2 или 13 (о разнице между ними будет сказано далее), конденсатор C2 фильтра НЧ – между выводом 9 и общим проводом.
Вывод 10 — выход усилителя-повторителя. Напряжение на нем в процессе работы микросхемы — это напряжение рассогласования, подаваемое на ФНЧ. Вывод 10 предназначен для того, чтобы напряжение рассогласования можно было при необходимости просто выделить без шунтирующего воздействия на конденсатор ФНЧ. К этому выводу допускается подключить резистор сопротивлением более 10 кОм.
Вывод 10 предназначен для того, чтобы напряжение рассогласования можно было при необходимости просто выделить без шунтирующего воздействия на конденсатор ФНЧ. К этому выводу допускается подключить резистор сопротивлением более 10 кОм.
Вывод 15 — на нем находится катод встроенного стабилитрона с напряжением стабилизации 5,6 вольт (напряжение стабилизации этого стабилитрона может быть иным, что зависит от производителя микросхемы). Данный стабилитрон можно при желании использовать в цепи питания микросхемы.
Вывод 16 — плюс питания микросхемы.
Входы и выходы фазовых компараторов ФК1 и ФК2
Меандр с выхода ГУН берется с вывода 4 и подается на вывод 3, присоединенный через усилитель-формирователь к входам фазовых компараторов ФК1 и ФК2. При желании сигнал с ГУН можно дополнительно пропустить через делитель частоты.
Вход 14 — сигнальный, на него и подается входной сигнал, с которым необходимо синхронизировать выходной сигнал на выходе ГУН. В зависимости от характера входного сигнала, разработчик может выбрать, какой из фазовых компараторов использовать: ФК1 или ФК2, и к выбранному компаратору присоединить резистор ФНЧ (к выводу 2 или 13). У фазового компаратора ФК2 есть индикаторный вывод 1, на нем появляется напряжение высокого уровня, когда сигналы максимально синхронизированы.
У фазового компаратора ФК2 есть индикаторный вывод 1, на нем появляется напряжение высокого уровня, когда сигналы максимально синхронизированы.
Особенность ФК1 в том, что он представляет собой простой логический элемент «исключающее ИЛИ», и качество его работы зависит от параметров ФНЧ на его выходе. Работа начинается с центральной частоты f0=(fmax-fmin)/2, имеется возможность захвата гармоники центральной частоты. Обладает высокой помехоустойчивостью.
Особенность ФК2 в том, что он обрабатывает лишь положительные перепады подаваемых на него импульсов, и скважность импульсов поэтому не имеет значения. Работа начинается с минимальной частоты fmin, возможности захвата гармоники центральной частоты нет. Обладает низкой помехоустойчивостью. В ФНЧ требуется конденсатор с малым током утечки. ФК2 лучше подходит для использования в силовых схемах с LC-резонансом.
Выбор навесных компонентов
В качестве фильтра низкой частоты ФНЧ устанавливаются резистор R3 и конденсатор C2. Для корректной работы ФАПЧ, постоянна времени RC должна быть в десятки раз больше примерной частоты захвата ФАПЧ.
Для корректной работы ФАПЧ, постоянна времени RC должна быть в десятки раз больше примерной частоты захвата ФАПЧ.
Как правило, частота захвата приблизительно известна разработчику, поэтому изначально задаются диапазоном автоподстройки частоты: fmin и fmax. По первой номограмме определяют, с учетом напряжения питания микросхемы и требуемого fmin, величины R2 и С1. Затем, по второй номограмме, исходя из требуемого соотношения fmax/fmin подбирают R1. Лучше предусмотреть возможность регулировки резисторов в схеме.
Ранее ЭлектроВести писали, как регулировать яркости светодиодов.
По материалам: electrik.info.
|
Основные принципы работы электронной памяти
Заметьте, что в начале идет условное обозначения типа микросхемы, а в конце указывается максимальная тактовая частота шины, на которой они могут работать. Объем памяти в микросхеме указывается в двух вариантах: 256Mb — общее количество ячеек памяти в микросхеме; 32Мх8 — это обозначение показывает, что на каждый разряд приходится по 32 Мбайт (также используется термин «глубина адресного пространства», от англ, address depth). Если умножить 32 Мбайт на 8, то получается 256 Мбайт.
|
|

 Но поскольку для управления триггером
требуются цепи управления, то элементарная запоминающая ячейка современной статической
памяти, которая применяется, в частности, для кэш-памяти, содержит иногда до
десятка транзисторов. Для примера на рис. 3.5 показана схема ячейки памяти КМОП-микросхемы.
В ней из шести КМОП-транзисторов только транзисторы V3 и V5 отвечают за хранение
информации, а остальные используются по другому назначению.
Но поскольку для управления триггером
требуются цепи управления, то элементарная запоминающая ячейка современной статической
памяти, которая применяется, в частности, для кэш-памяти, содержит иногда до
десятка транзисторов. Для примера на рис. 3.5 показана схема ячейки памяти КМОП-микросхемы.
В ней из шести КМОП-транзисторов только транзисторы V3 и V5 отвечают за хранение
информации, а остальные используются по другому назначению.


КОМБИНИРОВАННЫЕ МИКРОСХЕМЫ (ШИМ + КЛЮЧ
Значительно упростить процесс разработки и изготовления ИИП стало возможным благодаря появлению нового поколения ИМС, совмещающих в одном корпусе ШИМ, цепи управления и защиты, высоковольтный полевой транзистор. Такая комбинация устройств предельно сокращает количество компонентов схемы (простейший обратноходовый ИИП содержит 15 — 20 элементов), на 50% снижает габариты и массу устройства. При этом ИИП обладает высокой степенью защиты от перегрева и перегрузки, обладает хорошей электромагнитной совместимостью с другими устройствами. Стоимость комбинированных микросхем сопоставима со стоимостью мощных полевых транзисторов, применяемых в настоящее время в качестве выходных в ИИП. Если учесть экономию за счёт меньшего числа внешних компонентов, меньшего числа операций по сборке и наладке таких источников питания, то выгоды в промышленном производстве по сравнению с линейными источниками питания, ИИП на дискретных элементах и ШИМ, описанных ранее, становятся очевидными. Кроме невысокой стоимости ИИП на комбинированных микросхемах изготовители гарантируют конечному пользователю экономию за счёт меньшего потребления энергии самим устройством. В настоящее время на отечественном рынке представлены комбинированные микросхемы для ИИП производства Power Integrations (семейства TOPSwitch и последующих) и производства STMicroelectronics (семейства VIPer).
Такая комбинация устройств предельно сокращает количество компонентов схемы (простейший обратноходовый ИИП содержит 15 — 20 элементов), на 50% снижает габариты и массу устройства. При этом ИИП обладает высокой степенью защиты от перегрева и перегрузки, обладает хорошей электромагнитной совместимостью с другими устройствами. Стоимость комбинированных микросхем сопоставима со стоимостью мощных полевых транзисторов, применяемых в настоящее время в качестве выходных в ИИП. Если учесть экономию за счёт меньшего числа внешних компонентов, меньшего числа операций по сборке и наладке таких источников питания, то выгоды в промышленном производстве по сравнению с линейными источниками питания, ИИП на дискретных элементах и ШИМ, описанных ранее, становятся очевидными. Кроме невысокой стоимости ИИП на комбинированных микросхемах изготовители гарантируют конечному пользователю экономию за счёт меньшего потребления энергии самим устройством. В настоящее время на отечественном рынке представлены комбинированные микросхемы для ИИП производства Power Integrations (семейства TOPSwitch и последующих) и производства STMicroelectronics (семейства VIPer). Данные для выбора типа микросхемы в зависимости от требуемой мощности приведены в таблице.
Данные для выбора типа микросхемы в зависимости от требуемой мощности приведены в таблице. Принципы работы ШИМ комбинированных микросхем не отличаются от применяемых в ИМС, описанных ранее, они содержат те же основные узлы. Принципиальное отличие заключается в исполнении на одном кристалле и оформлении в одном корпусе (имеющем для разных типов микросхем от 3 до 8 выводов) всех цепей ИИП, кроме входных выпрямителей/фильтров, импульсного трансформатора, вторичных цепей, цепей обратной связи, конденсатора вывода управления. Для того, чтобы была возможность полностью оценить простоту реализации ИИП на комбинированных микросхемах, на рисунке приведена типовая схема включения ИМС семейства TOPSwitch, которое является одним из первых в этом классе. Типовые схемы других семейств выполнены примерно также, с тем же количеством элементов, но с большим числом функций.
Типовая схема ИИП для работы от сети переменного тока на ИМС семейства TOPSwitch:
Основные параметры комбинированных ИМС для импульсных источников питания:
| Семейство | Тип ИМС | Типы корпусов | Рекомендуемый диапазон мощности (Вт) | Особенности применения | Uраб, В1 | Диапазон рабочих температур, °С2 | Fраб, кГц | Рабочий цикл, % | Дополнительные функции | |||||||
Защита от пониж. Uпит Uпит |
Защита от повыш. Uпит | Дежурный режим | «Мягкий» пуск | Внешнее ограничение тока | ЧМ рабочей частоты | Внешняя синхронизация | ДУ вкл/выкл | |||||||||
| TOPSwitch | TOP200… 204, 214 |
TO-220/3 | 0…100 (в схемах корректора фактора мощности до 150) | 36…700 | -40…+145 | 100+10 | 1,8…67 | + | ||||||||
| TOP100…104 | 0…60 (в схемах корректора фактора мощности до 110) | Для работы в сети 110/100 В | + | |||||||||||||
| TOP209 | DIP8 | 0…8 | 36…700 | 100+10 (TOP210) 70+15 (TOP209) |
+ | + | ||||||||||
| TOPSwitchII | TOP221…227 | TO220, PDIP8, SMD8 | 0…150 | -40…+135 | 100+10 | 1,7…67 | + | + | ||||||||
| TOPSwitch-FX | TOP232…234 | TO220-7B, PDIP8, SMD8 | 0…75 | Возможна работа на частоте 0,5Fраб для уменьшения помех | 132+8 или 66+4,5 | 1,5…78 | + | + | + | + | + | + | + | + | ||
| TOPSwitch-GX | TOP242…250 | TO220-7B, PDIP8, SMD8, TO263-7C, TO262-7C | 0…290 | -40…+140 | 03…66,8 | + | + | + | + | + | + | + | + | |||
| TinySwitch | TNY253…255 | DIP8, SO8 | 0…10 | Возможна работа без обмотки ОС импульсного трансформ. (Типовая схема включения) (Типовая схема включения) |
50…700 | -40…+135 | 44+4 (TNY253, 254) 130+15 (TNY255) | 03…68 | + | + | + | |||||
| TinySwitchII | TNY256 | TO220-7B, DIP8B, SOIC8B | 0…23 | 130+15 | 03…66 | + | + | + | + | |||||||
| TinySwitch Plus | TNY264, 266…268 | 0…19 | 132+8 | 03…65 | + | + | + | + | ||||||||
| DPA-Switch | DPA423…426 | TO263-7C | 0…83 (max 100) | Для построения DC-DC конвертеров со входным напряжением 36…75 В | 16…220 | -40…+137 | 400+25 или 300+18 | 03…75 | + | + | + | + | + | + | + | |
| LinkSwitch | LNK500, 501 | DIP8B, SOIC8 | 0…5,5 | Для построения адаптеров и зарядных устройств | 50…700 | -40…+135 | 42+7,5 (LNK500) 42+4 (LNK501) | 1,8…77 | + | + | ||||||
| VIPer | VIPer20/50/100 | TO220/5, DIP8 | До 20/50/100 | До 620, до 700 (с индексами A, ASP) | -40…+170 | 100+10, возможна работа до 200 кГц | 03…65 | + | + | + | + | + | ||||
Примечание:
1) В качестве нижнего предела указано напряжение стока, при котором гарантируется работа ИМС с указанными параметрами. Возможна работа при более низких напряжениях, но с отклонением параметров от нормы. В качестве верхнего предела указано напряжение пробоя исток-сток выходного транзистора.
Возможна работа при более низких напряжениях, но с отклонением параметров от нормы. В качестве верхнего предела указано напряжение пробоя исток-сток выходного транзистора.
2) Для всех микросхем (кроме семейства VIPer) указан рабочий диапазон температур -40:+150°С, однако в таблице в качестве верхней границы приведена температура срабатывания термозащиты.
3) У ИМС данных типов при отключении нагрузки уменьшается рабочий цикл (за счет пропуска рабочих периодов), а у некоторых — и рабочая частота. В результате в этих условиях рабочий цикл снижается практически до 0.
Коротко о некоторых функциях комбинированных микросхем. Все описываемые микросхемы имеют встроенные цепи авторестарта (защищают ИИП и нагрузку в случае аварии — КЗ нагрузки, обрыв петли ОС), ограничения тока стока (защита выходного транзистора), цепи запуска при подаче напряжения питания (снижается количество внешних компонентов), термозащиты. У микросхем семейств TOPSwitch и TOPSwitchII термозащита выполнена с внутренней защёлкой (после перегрева необходим перезапуск устройства), у остальных — с гистерезисом температуры срабатывания (после остывания происходит автоматический перезапуск). Все ИМС имеют возможность внешней блокировки работы выходного каскада. Наличие таких цепей значительно снижает вероятность выхода из строя ИИП на комбинированных микросхемах.
Все ИМС имеют возможность внешней блокировки работы выходного каскада. Наличие таких цепей значительно снижает вероятность выхода из строя ИИП на комбинированных микросхемах.
Для ИИП на основе ИМС семейств TOPSwitch и TOPSwitchII в некоторых условиях может потребоваться подключение искусственной нагрузки. Для остальных микросхем за счет усовершенствованных схемных решений это не требуется — рабочий цикл на холостом ходу снижается практически до 0.
Частотная модуляция рабочей частоты преобразователя снижает уровень побочных излучений на 5:10 дБ, что улучшает электромагнитную совместимость устройств. Возможность внешней синхронизации предусматривает синхронизацию от внешнего источника с частотой ниже, чем частота внутреннего генератора ИМС.
Наличие режима ДУ делает простой реализацию ИИП с микроконтроллерным управлением.
Наименование
К продаже
Цена от
К продаже:
5 235 шт.
К продаже:
7 032 шт.К продаже:
1 513 шт.
К продаже:
10 234 шт.К продаже:
479 шт.
К продаже:
398 шт.К продаже:
112 156 шт.
К продаже:
201 шт.К продаже:
1 266 шт.
К продаже:
50 шт.К продаже:
408 шт.
К продаже:
112 шт.К продаже:
43 шт.
К продаже:
56 шт.К продаже:
482 шт.
К продаже:
219 шт.К продаже:
618 шт.
К продаже:
1 164 шт.К продаже:
902 шт.
К продаже:
35 шт.К продаже:
8 шт.
К продаже:
1 480 шт.К продаже:
188 шт.
К продаже:
137 шт.К продаже:
34 939 шт.
К продаже:
8 447 шт.К продаже:
63 шт.
К продаже:
846 шт.К продаже:
200 шт.
К продаже:
1 661 шт.К продаже:
272 шт.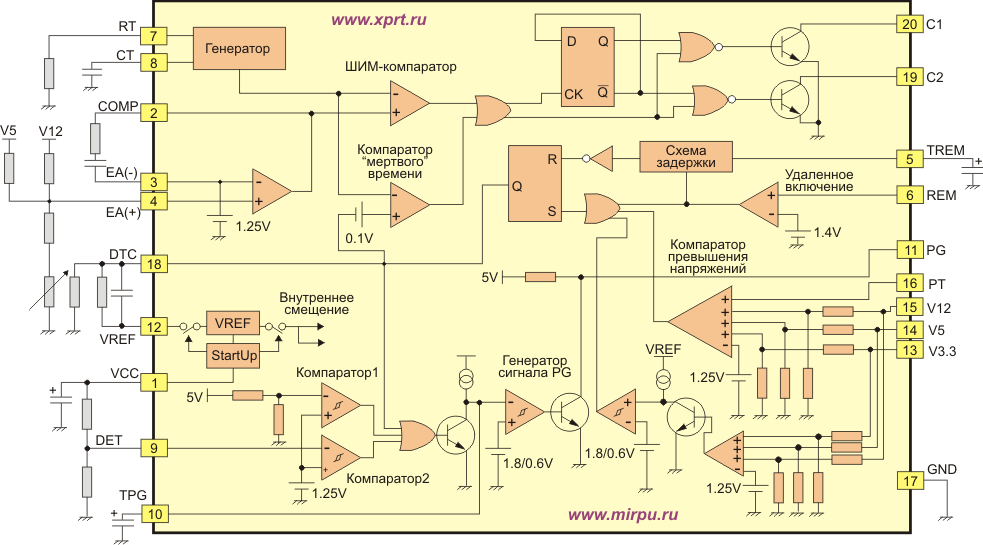
К продаже:
3 285 шт.К продаже:
3 348 шт.
К продаже:
1 967 шт.К продаже:
3 505 шт.
К продаже:
3 948 шт.К продаже:
674 шт.
К продаже:
797 шт.К продаже:
1 499 шт.
К продаже:
10 282 шт.К продаже:
101 шт.
К продаже:
244 шт.К продаже:
390 шт.
К продаже:
110 шт.К продаже:
1 000 шт.
К продаже:
19 272 шт.К продаже:
27 шт.
К продаже:
1 539 шт.К продаже:
8 741 шт.
К продаже:
1 105 шт.К продаже:
136 шт.
К продаже:
57 шт.К продаже:
352 шт.
К продаже:
4 095 шт.К продаже:
3 шт.
К продаже:
7 432 шт.К продаже:
230 шт.
К продаже:
171 шт.К продаже:
255 шт.
К продаже:
2 026 шт.К продаже:
150 шт.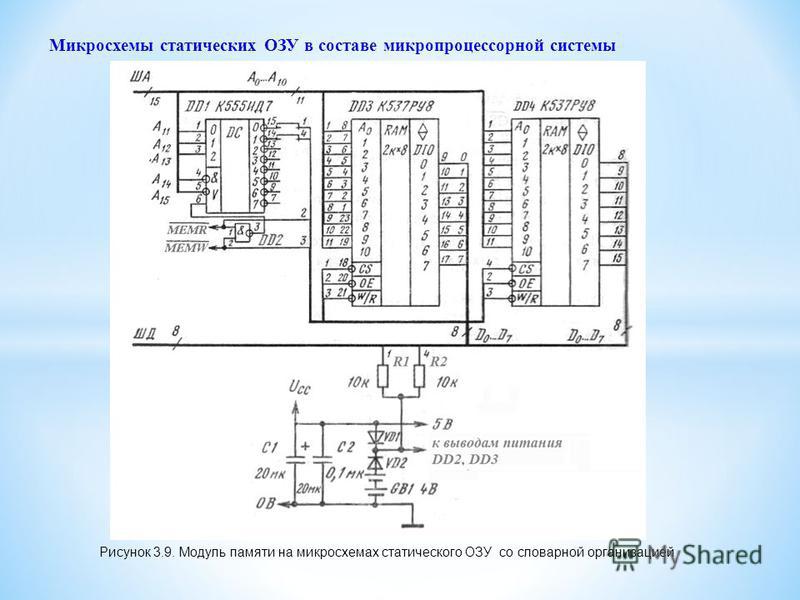
К продаже:
7 шт.К продаже:
215 шт.
К продаже:
2 128 шт.К продаже:
284 шт.
К продаже:
2 041 шт.К продаже:
941 шт.
К продаже:
64 шт.К продаже:
51 шт.
К продаже:
1 609 шт.К продаже:
98 шт.
К продаже:
298 шт.К продаже:
101 шт.
К продаже:
451 шт.К продаже:
10 шт.
К продаже:
122 шт.К продаже:
627 шт.
К продаже:
300 шт.К продаже:
92 шт.
К продаже:
905 шт.К продаже:
53 шт.
К продаже:
5 000 шт.К продаже:
5 482 шт.
К продаже:
69 048 шт.К продаже:
1 500 шт.
К продаже:
24 883 шт.К продаже:
3 000 шт.
К продаже:
42 шт.К продаже:
920 шт.
К продаже:
2 918 шт.К продаже:
5 590 шт.К продаже:
4 488 шт.К продаже:
700 шт.К продаже:
2 045 шт.К продаже:
899 шт.К продаже:
620 шт.К продаже:
72 шт.К продаже:
54 шт.К продаже:
213 шт.К продаже:
18 280 шт.К продаже:
27 шт.К продаже:
298 шт.К продаже:
1 021 шт.К продаже:
104 шт.К продаже:
20 шт.К продаже:
44 шт.К продаже:
1 911 шт.К продаже:
171 шт.К продаже:
2 668 шт.К продаже:
3 335 шт.Забота о микросхемах | Природа неврологии
Микросхемы: интерфейс между нейронами и функцией глобального мозга
- S Grillner &
- A M Graybiel
The MIT Press, 2006 г. 472 стр., Твердая обложка, 55,00 долл. США ISBN 0262072785 | ISBN: 0-262-07278-5
Поиск в Интернете по запросу «микросхемы» дает более полумиллиона результатов, и эта книга возглавляет список.К чему весь такой ажиотаж? Одна из причин заключается в том, что большинство нейробиологов изучают микросхемы в широком смысле слова, что делает этот том интересным. Редакторы дают гибкое рабочее определение (« четко определенные, довольно маленькие образования нервных клеток ») и обрисовывают в общих чертах свою цель — обеспечить мост от генетического и молекулярного уровня к поведенческому и когнитивному уровню не напрямую, а через понимание промежуточных микросхем. .
Книга состоит из четырех разделов, в которых рассматриваются микросхемы в двигательной системе, полосатом теле, обонятельной системе и неокортексе. Книга состоит из глав, написанных ведущими экспертами, за которыми следует групповой отчет.В первом разделе рассматриваются колликулярные микросхемы для саккадических движений глаз (Isa и Sparks), а также микросхемы локомоторных позвоночных (Sillar и Grillner) и беспозвоночных (Pfluger и Buschges) и подчеркивается тот факт, что исследователи добились большого прогресса в достижении этой общей цели и что исследователи в совершенно разных системах говорят на одном языке. Групповой отчет (Kiehn) представляет собой элегантную попытку извлечь общие принципы проектирования в центральных микросхемах генератора шаблонов в моторных системах, которые адаптированы для многих конкретных задач.Аргумент, эффективно поддерживаемый схемотехникой, состоит в том, что центральные генераторы паттернов включают почти универсальное «ядро», которое состоит из небольшой сети возбуждающих и тормозящих нейронов. Затем базовая микросхема функционально адаптируется к конкретным задачам (например, сегментарно реплицируется и соединяется между собой в опорно-двигательных системах или с помощью кардиостимулятора в дыхательных центрах ствола мозга). Нейромодуляция важна для настройки выхода схем.
Эти темы развиваются, и новые вводятся в раздел полосатого тела, который знакомит читателей с основными типами клеток, микросхемами и нейромодуляцией в базальных ганглиях (Surmeier, Tepper and Plenz, Bergman, Kimura, Wickens).В групповом отчете (Bolam) обсуждается каноническая микросхема, взаимосвязанная группа нейронов со средними шипами, ГАМКергические интернейроны с быстрым выбросом и гигантский холинергический интернейрон, коллективно иннервируемый кортикостриатными глутаматергическими проекциями и дофаминергическими проекциями черной субстанции. Эта архитектура предлагается в качестве субстрата для «выбора» между различными корковыми входами с помощью хорошо охарактеризованных ионных механизмов, опосредующих состояния «ВВЕРХ» и «ВНИЗ». Допаминергическая и холинергическая модуляция, действующая на каноническую микросхему в сильно дивергентной и конвергентной системе корковых афферентов, предлагается в качестве основы для пластичности, связанной с вознаграждением.
В разделе об обонятельных микросхемах пересматриваются общие принципы сенсорного кодирования и декодирования систем беспозвоночных и позвоночных. В главах рассматриваются популяционное кодирование и сетевая динамика в обонятельных путях насекомых (Laurent), замена нейронов в обонятельной луковице (Lledo), молекулярная спецификация связи в обонятельных микросхемах (Mombaerts, Feinstein) и топографические аспекты обонятельных цепей, карт и кодов (Sachse, Galizia). ). Групповой отчет (Фридрих) фокусируется на формулировании канонического взгляда на обонятельные микросхемы, включая пространственные и временные аспекты.
Неокортикальный отдел широко варьируется, включая разнообразие интернейронов (DeFilipe et al .), Состояния UP коры, цепочки синфайров и сетевую динамику (McCormick и Yuste), динамику всего мозга, относящуюся к сознанию (Changeux и Michel), и вычислительную подходы (Маасс и Маркрам). В групповом отчете (Fregnac) исследуется степень каноничности корковых цепей в разных областях и видах и связь UP-состояний с корковыми микросостояниями. В отчете оценивается модульность (непрерывная или кластерная организация внутри и между корковыми областями), подчеркивая вертикальный и горизонтальный характер столбчатой организации.В отчете также рассматриваются временные аспекты обработки информации и «шума» в корковых цепях: регулировка усиления, периодическое возбуждение и многое другое. Группа смело рассматривает, как это может быть связано с тем, что делает неокортекс, находя достоинства механизмов микросхем (цепочек синфайров, состояний UP) во внимании, восприятии и сознании.
В книге кратко излагаются ключевые темы исследования микросхем: кодирование пространственной информации, временная обработка информации, возбуждающе-тормозные взаимодействия, модульность и модуляция.Многие главы освежающе спекулятивны, поднимаясь к цели серии конференций Далема — выйти за рамки формата большинства материалов конференций. Действительно, особая сила книги состоит в том, что многие авторы явно перечисляют то, что они считают ключевыми неотвеченными вопросами в своей подполе. Большая часть предположений основана на нейронной «основе» — экспериментально определенных ионных и синаптических механизмах, лежащих в основе организации цепей. В этой книге разъясняется, что поведение в конечном итоге следует понимать с точки зрения функциональной организации микросхем.
На будущее перед нами стоят два вопроса. Во-первых, необходимо уточнить уровень организации при идентификации и анализе микросхем. Первоначально применяемый к локальным паттернам синаптического взаимодействия и простым рефлекторным путям, этот термин теперь применяется на многих уровнях, включая внутридендритные взаимодействия, локальные синаптические связи, межслойные связи и локальные цепи. Другой вопрос, являются ли модули микросхем в коре головного мозга разнообразными или стереотипными, и как это связано с высшими кортикальными функциями.Многие рабочие гипотезы, изложенные в этом томе, будут стимулировать дальнейшие исследования таких вопросов.
Информация об авторе
Принадлежность
Гордон М. Г. Шеперд работает на кафедре физиологии Медицинской школы Файнберга Северо-Западного университета, Чикаго, Иллинойс 60611, США. [email protected]
Гордон М. Дж. Шеперд
Гордон М. Шеперд работает на кафедре нейробиологии Медицинской школы Йельского университета, 333 Cedar Street, New Haven, Connecticut 06510, [email protected]
Gordon M Shepherd
Об этой статье
Цитируйте эту статью
Shepherd, G., Shepherd, G. Обращение к микросхемам. Nat Neurosci 10, 1503 (2007). https://doi.org/10.1038/nn1207-1503
Ссылка для скачивания
Модель индивидуализированных канонических микросхем, поддерживающих когнитивные операции
Abstract
Считается, что основные когнитивные функции, такие как язык, память и принятие решений, зависят от распределенных сетей из большого количества базовых элементов, называемых каноническими микросхемами.В этом теоретическом исследовании мы предлагаем новую каноническую модель микросхемы и обнаруживаем, что она поддерживает две основные вычислительные операции: стробирующий механизм и рабочую память. С помощью бифуркационного анализа мы систематически исследуем динамическое поведение канонической микросхемы в отношении параметров, которые управляют балансом локальной сети, то есть отношения между возбуждением и торможением и ключевыми внутренними архитектурами обратной связи канонических микросхем. Мы связываем локальное поведение канонической микросхемы с когнитивной обработкой и демонстрируем, как сеть взаимодействующих канонических микросхем позволяет устанавливать пространственно-временные последовательности в контексте синтаксического анализа во время понимания предложения.Это исследование обеспечивает основу для использования индивидуализированных канонических микросхем для построения биологически реалистичных сетей, поддерживающих когнитивные операции.
Образец цитирования: Kunze T, Peterson ADH, Haueisen J, Knösche TR (2017) Модель индивидуализированных канонических микросхем, поддерживающих когнитивные операции. PLoS ONE 12 (12): e0188003. https://doi.org/10.1371/journal.pone.0188003
Редактор: Томас Веннекерс, Плимутский университет, СОЕДИНЕННОЕ КОРОЛЕВСТВО
Поступила: 29 мая 2017 г .; Одобрена: 25 октября 2017 г .; Опубликован: 4 декабря 2017 г.
Авторские права: © 2017 Kunze et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все соответствующие данные находятся в документе и его файлах с вспомогательной информацией. Исходный код доступен по адресу https://github.com/TiKunze/CanMics.
Финансирование: Эта работа финансировалась Обществом Макса Планка и поддерживалась программой совместных исследований DAAD-Go8-Germany в рамках гранта номер 57061157.A.D.H.P была поддержана исследовательским сообществом сэра Джона Эклза. J.H. получил частичное финансирование от Министерства науки Тюрингии в рамках гранта № 2015 FGR 0085.
Конкурирующие интересы: Авторы заявили, что конкурирующих интересов не существует.
Введение
Большинство современных нейробиологических теорий основаны на коннекционистском подходе, в котором высшие когнитивные функции закреплены в распределенной сети из большого количества схожих базовых элементов, часто называемых каноническими микросхемами [1–7].Эти относительно простые элементы усложняют когнитивную обработку в силу (i) их взаимодействия в большом количестве в рамках организованной сетевой топологии и (ii) индивидуальной настройки их свойств. Хотя более высокие когнитивные операции обязательно связаны с объединением и распределением больших объемов информации и, следовательно, должны полагаться на соединительную структуру более широкой сети, канонические микросхемы могут играть важную роль, предоставляя набор из базовых операций [2, 8 , 9].В этом исследовании мы предлагаем общую вычислительную структуру для кортикальной канонической микросхемы. Мы систематически исследуем способность этой модели представлять важные базовые операции, количественно оценивать влияние фундаментальных структурных особенностей и физиологических переменных и демонстрируем ее способность сотрудничать в более крупных сетях для реализации когнитивных функций.
Две из наиболее фундаментальных базовых операций на локальном уровне — это стробирование потока сигналов и рабочая память .Стробирование потока сигналов контролирует передачу нейронных сигналов. Вероятно, это будет зависеть (i) снизу-вверх от свойств, в частности, значимости, самого входного сигнала и (ii) от нисходящей модуляции канонической микросхемы глобальной сетью. Выбор ввода в соответствии с его значимостью, то есть его значимостью с точки зрения величины и продолжительности, например, связан с управлением зрительным вниманием [10], избирательной реакцией на сенсорный ввод и определением путей обработки [11].
Для обработки временно структурированной информации требуется механизм быстрой рабочей памяти, который не полагается на структурные (например, синаптические) изменения [12]. Бистабильная динамика в канонической микросхеме — одна из возможных реализаций такого механизма.
В отличие от других частей мозга, таких как ствол мозга, мозжечок или таламус, схема коры в основном характеризуется повторяющимися возбуждающими и тормозящими петлями обратной связи на локальном уровне и двунаправленной разреженной возбуждающей связностью на глобальном уровне [ 13].Таким образом, модель локальной корковой микросхемы должна включать пирамидные клетки с длинными аксонами, выступающими в отдаленные области коры, а также локальные возбуждающие и тормозные петли обратной связи. Такие базовые архитектуры, обнаруженные в различных пространственных масштабах, были представлены в скупой форме моделей нейронной массы / поля [14–18]. Исследования стационарного поведения таких моделей показали, что они действительно могут обеспечить основу для вышеупомянутых основных операций, показывая бистабильность и бифуркации [19–21].Здесь мы распространяем эти результаты на реакции на кратковременные стимулы и тем самым оцениваем реализацию стробирования сигнала и операций с рабочей памятью. В этом контексте мы исследуем влияние следующих структурных и физиологических проблем, которые ранее не изучались в моделях нейронной массы канонических микросхем:
- Непрямая возбуждающая обратная связь в сравнении с прямой. Большинство моделей нейронной массы и поля рассматривают только одну возбуждающую нейронную популяцию [e.ж., 17], где петля возбуждающей обратной связи моделируется как рекуррентная (прямая) обратная связь. Другие подходы [например, 16] различают пирамидные ячейки, которые обеспечивают выходную связь на большие расстояния, и возбуждающие интернейроны, которые являются основными приемниками восходящего входа [2, 22]. В этих трехпопуляционных моделях возбуждающая обратная связь с пирамидными клетками является непрямой, опосредованной возбуждающими интернейронами. Гарнье и его коллеги [23] исследовали последствия прямой и косвенной возбуждающей обратной связи и сообщили, что путь косвенной обратной связи обеспечивает дополнительную динамику.Однако уместность этой дополнительной динамики должна быть сбалансирована с затратами на увеличение сложности модели и должна быть оценена с учетом конкретных требований моделирования [2]. Поэтому оценим чувствительность основных операций канонической микросхемы по отношению к этому выбору.
- Повторяющаяся тормозящая обратная связь. Аксоны тормозных интернейронов образуют коллатерали, нацеленные на те же или другие тормозные нейроны. Это «подавление торможения» включено в некоторые модели [17], в то время как в других оно не принимается во внимание [16].Нелинейный эффект этого растормаживания по отношению к основным операциям микросхемы будет исследован с помощью численного моделирования.
- Баланс локальной сети. Было показано, что взаимосвязь между торможением и возбуждением в нейронной сборке имеет центральное значение для ее способности обрабатывать информацию [24]. Как следствие, он опосредует функции мозга более высокого порядка [25], а его нарушение нарушает механизмы обработки коры головного мозга и может привести к серьезным сбоям и расстройствам мозга, таким как эпилепсия [26, 27], аутизм [28-30], шизофрения [31, 32] и эксайтотоксичность [33].Здоровый мозг автоматически устанавливает динамический баланс возбуждения и торможения. Это было показано теоретически [34] и экспериментально в исследованиях in vitro, [35] и in vivo, [26, 36]. Поэтому в рамках этого исследования изучается, как баланс локальной сети влияет на функциональные возможности локальных микросхем и как можно индивидуализировать канонические микросхемы.
Сети, состоящие из множества канонических микросхем, реализованных с помощью моделей нейронной массы, недавно были использованы для объяснения экспериментальных данных в нейрокогнитивных экспериментах, особенно в рамках динамического каузального моделирования (DCM) [37, 38].Однако мало внимания уделяется двум аспектам: (i) взаимосвязи между сетевым поведением и внутренними свойствами микросхем и (ii) механистическим объяснением поведения, а не данными изображений мозга. Здесь мы описываем в качестве простого примера сеть обработки предложений, состоящую из канонических микросхем, которая гибко обрабатывает расположение слов и позволяет различать альтернативные интерпретации неоднозначных предложений. Мы показываем, что точная настройка баланса локальной сети имеет решающее значение для функционирования предлагаемой модели обработки предложений.
Методы
Описание канонической модели нейронной популяции
Далее мы представляем модель нейронной массы, которая используется в этом исследовании. Модели нейронной массы — это устоявшийся подход к объяснению данных электроэнцефалографии [16, 38, 39], выяснения эпилептогенных процессов [40, 41] и электрической стимуляции мозга [42, 43], а также исследования динамического поведения ограниченной нервной области [19–19]. 21]. В этом исследовании мы используем модель нервной массы, которая имеет три нейронные массы или популяции, представляющие пирамидные клетки (Py), возбуждающие интернейроны (EIN) и тормозные интернейроны (IIN).Две популяции интернейронов образуют петли обратной связи на Py (Fig 1A). Каждая из этих нейронных масс описывается средним мембранным потенциалом V (t), который связан со средней скоростью возбуждения φ (t) популяции через нелинейную функцию активации S (V (t)). Для определения и параметризации синаптического ответа и функций активации мы следуем подходу Шпиглера [20], который основан на более ранних описаниях [16, 39]. В каждой нервной массе афферентная средняя частота возбуждения φ (t), достигающая дендритного дерева нейронной популяции, преобразуется в соответствующий средний мембранный потенциал V (t) путем свертки частоты возбуждения с ядром синаптического ответа h e, i (t) как в (1) где индекс e ( i ) обозначает ядро синаптического ответа возбуждающей (тормозной) нервной массы.Ядро синаптического ответа моделируется как альфа-функция. (2) где θ (t) обозначает функцию Хевисайда, H e, i — синаптическое усиление, отражающее количество и эффективность синаптических контактов, а τ e , i — характерная постоянная времени любого возбуждающего или тормозящие действующие нервные массы. Тогда средний мембранный потенциал V c (t), c ∈ [P, E, I] соответствующих нейронных масс зависит от суммы всех входящих компонентов.Используя функцию Грина, это можно выразить как: (3) где D — временной дифференциальный оператор второго порядка, который имеет вид (4) где уравнение (2) представляет собой функцию Грина этого дифференциального оператора. Затем этот оператор разделяется на два дифференциальных уравнения первого порядка. Преобразование среднего мембранного потенциала в среднюю частоту возбуждения, представляющую процессы, происходящие на аксональном бугорке нейрона, моделируются сигмоидальной функцией активации, в данном случае логистической функцией (5)
Рис 1.Обобщенная архитектура модели нейронной массы.
A) Модель нервной массы учитывает возбуждающие интернейроны (EIN), тормозящие интернейроны (IIN) и пирамидные клетки (Py). Параметр архитектуры b 1 управляет развертыванием прямой и непрямой возбуждающей обратной связи, а также популяцией, принимающей вход, тогда как учет тормозящих коллатералей регулируется параметром архитектуры b 2 . Эта параметризация позволяет провести сравнительное исследование соответствующих изменений в динамическом поведении трех различных архитектур: B) модель трех популяций, C) модель двух популяций и D) модель двух популяций с повторяющейся тормозящей обратной связью. ИИН.Передаваемые средние скорости возбуждения φ (t) масштабируются с помощью выигрыша в связности N ab между исходной совокупностью, b , и целевой совокупностью, a , соответственно. Мембранный потенциал пирамидных клеток, V py (t) = V 2 (t) -V 3 (t), представляет собой результат модели (обозначен красными стрелками), который можно обнаружить, например, по ЭЭГ.
https://doi.org/10.1371/journal.pone.0188003.g001
В то время как e 0 представляет половину наивысшей достижимой скорости стрельбы, r — максимальный наклон сигмовидной функции и v 0 обозначает мембранный потенциал, для которого задействована половина максимальной скорости воспламенения.Средний мембранный потенциал Py, объединяющий как положительную, так и отрицательную обратную связь, формирует наблюдаемый сигнал цепи (например, посредством ЭЭГ) и, в то же время, вызывает выходной сигнал в отдаленные области через функцию активации. Следовательно, описание наблюдаемой динамики сосредоточено на этой основной клеточной популяции.
Мы строим общий формализм модели, который учитывает различные топологии локальной обратной связи: (i) трехпопуляционная модель с косвенным путем возбуждающей обратной связи через EIN (рис. 1B), (ii) двухпопуляционная модель с прямой возбуждающей обратной связью через самосоединения Py (рис. 1C) и (iii) двухпопуляционная модель с прямой возбуждающей обратной связью и повторяющейся тормозной обратной связью IIN (рис. 1D).Локальные топологии управляются двумя параметрами: b 1 и b 2 . Первый параметр (b 1 ) допускает постепенный переход между моделями с двумя популяциями с внешним входом p ext , полученным возбуждающим населением (Py), и трехпопуляционной моделью с внешним входом, p доб , полученный возбуждающими интернейронами (EIN). Важно, что можно смоделировать промежуточные ситуации, когда обе возбуждающие популяции получают внешнюю информацию и сосуществуют как прямые, так и непрямые петли возбуждающей обратной связи (0 1 <1).Этот подход позволяет рассматривать выбор принципа в структуре модели (модель с двумя или тремя популяциями) как непрерывный параметр, который может быть подвергнут, например, бифуркационному анализу. См. Файл S1 для получения более подробной информации о сопоставлении трех моделей с двумя популяциями. Второй параметр (b 2 ) контролирует наличие рекуррентной петли обратной связи для IIN. Согласно схеме, изображенной на рис. 1А, система управляющих уравнений канонической микросхемы имеет следующий вид: (6)
Параметры N ab обозначают прирост связности между исходной популяцией b и целевой популяцией a , где a , b ∈ [P, E, I].Для численного интегрирования этой системы нелинейно связанных линейных обыкновенных дифференциальных уравнений используется метод Гойна. Операторы D e , i и S (∙) обозначают возбуждающий (тормозной) оператор временного дифференциала и сигмоидальную функцию активации, соответственно (см. Уравнения (4) и (5)). Была проверена стабильность интегрирования на интервале интегрирования 1 мс. Система изначально параметризована в соответствии с ранее использованной конфигурацией [16, 39], см. Таблицу 1.Кроме того, прирост связности N PP и N II определяется как N PP = 113,4 (см. Файл S1) и N II = 33,25, аналогично другим силам подавления соединения.
Определение и параметризация баланса сети
Взаимосвязь торможения и возбуждения, часто называемая сетевым балансом, в нейронной сборке регулирует взаимодействие нейронных единиц, влияет на динамику состояний мозга и связана с тяжелыми мозговыми расстройствами, такими как эпилепсия [26, 27], аутизм [28–30] или шизофрения [31, 32].Концепция сетевого баланса неоднозначна, и ее трудно измерить просто как соотношение возбуждения и торможения в нейронной системе. Это связано с множеством пространственных и временных масштабов в мозге [24, 28] и множеством структурных и функциональных аспектов, которые можно было бы рассмотреть. Описание баланса сети на мезоскопическом уровне взаимодействующих нейронных популяций может быть сосредоточено на структурных влияниях, таких как топология, количество и эффективность синаптических контактов, или функциональных характеристиках, таких как передаваемая частота возбуждения или факторы синаптического ответа.В вычислительных моделях других исследований предложенные подходы связывают возбуждающие и тормозные заряды, проводимость [27, 28, 36] или мембранные потенциалы [24] друг с другом. Часто сетевой баланс определяется в сетевом контексте как отношение повторяющегося торможения к возбуждению [44, 45]. Однако это ограничено двумя моделями населения и становится неоднозначным для моделей нескольких популяций, таких как те, которые используются в нашем исследовании.
В нашей модели подходящие параметры, которые потенциально имеют отношение к возбуждению и торможению, включают: (i) функцию синаптического ответа (постоянные времени, синаптический выигрыш), (ii) внешний вход для всех трех популяций, (iii) параметры сигмоидальная активационная функция и (iv) усиление связности между популяциями.Среди этих параметров синаптические приросты H e, i , прирост связности, то есть N EP , N PE , N PI , N IP N PP и N II , и внешние воздействия оказывают наиболее прямое и биологически правдоподобное воздействие на возбуждение и торможение системы. Однако обратите внимание на формальную избыточность, но концептуальную разницу между синаптическими усилениями и связями в уравнениях системы. Согласно основным уравнениям модели, т.е.е. Уравнения (1) — (6), H i , N PI и N II отражают усиление тормозной обратной связи только при другом масштабировании. Кроме того, изменение H e эквивалентно синхронному изменению N EP , N PE , N IP и N PP . Таким образом, в интересах управляемости для исследования модулирующего влияния возбуждения и торможения на локальную динамику достаточно рассмотрения скупого набора параметров. Следовательно, мы фокусируем наш анализ на влиянии H e и H i , которые интерпретируются как отражающие эффективность и плотность возбуждающего вещества, т.е.г. AMPA и ингибиторы, например ГАМК А , рецепторы нейромедиаторов. Это эквивалентно количеству и силе синаптических весов.
Бифуркационный анализ, моделирование и карта динамических функций
Уравнения модели были смоделированы в безразмерной форме в Matlab (The MathWorks, Inc., Натик, Массачусетс, США), а бифуркационный анализ был выполнен с использованием инструмента численного продолжения DDE-BIFTOOL [46]. Были использованы стандартные методы для расчета кривых с фиксированной точкой, т.е.е. вычисление неподвижных точек, вывод матрицы Якоби, линеаризация системы вокруг неподвижных точек и оценка собственных значений для определения локальной устойчивости. Синаптические коэффициенты усиления H e, i служили параметрами бифуркации в соответствующих локальных топологиях. Продолжительность моделирования составляла 5 секунд, а переменные состояния инициализировались нулевым вектором. Из-за инициализации системы с внешним входным уровнем p ext = 0s -1 , система постоянно находилась на нижней ветви S-образной кривой с фиксированной точкой в случае бистабильного режима.
В каждом моделировании модель стимулировалась прямоугольным импульсом определенной интенсивности в диапазоне от 50 с -1 до 250 с -1 и длительностью от 500 до 1500 мс, начиная с времени установления 1 с. Поведение динамической реакции канонической микросхемы на эту стимуляцию было классифицировано в пределах трех различных временных окон путем сравнения максимальных мембранных потенциалов популяции пирамидных клеток с порогом срабатывания. Временными окнами были: i) окно предварительного стимула (0.5 с – 1 с), ii) окно немедленного отклика (от 1,1 до 3,5 с) и iii) асимптотическое окно (от 4 до 5 с), см. Рис. 2A. Порог срабатывания u th = 4 мВ был определен относительно максимальной скорости срабатывания 5 с -1 , так что на пороге достигается около 25% максимальной скорости срабатывания. В каждом временном окне система считалась активной, если максимальная активация превышала порог, и неактивной, если максимальная активация была ниже порога. Наблюдались три основных типа поведения (см. Рис. 2B): i) поведение памяти, когда система остается постоянно активной после прекращения ввода, ii) поведение передачи, когда активность выше порога во время окна немедленного ответа, но ниже в остальных случаях. и iii) невосприимчивое поведение, когда максимальная активность постоянно выше или ниже порогового значения во всех трех окнах.Обратите внимание, что в некоторых случаях активность колеблется около порогового значения. В этих случаях популяция может активировать постсинаптические популяции, по крайней мере, на некоторое время, и поэтому считается, что она превышает пороговую величину. Возникновение этого поведения в зависимости от управляющих параметров суммировано в так называемых картах динамических функций , которые типичны для репертуара динамических откликов соответствующей параметризации.
Рис. 2. Принцип стимуляции и категоризация поведения динамической реакции.
A) Модель получала прямоугольную стимуляцию различной интенсивности и продолжительности (зеленая линия). Максимум среднего мембранного потенциала популяции пирамидных клеток (синяя линия) регистрировали в трех временных окнах, то есть в окне предварительного стимула, окне немедленного ответа и асимптотическом окне (серые заштрихованные области). Для классификации поведения реакции эти значения активации сравнивались с порогом 4 мВ (красная горизонтальная линия, u -й ) в каждом окне — «0» обозначает подпороговую активацию, а «1» обозначает активацию, превышающую пороговое значение.Б) Комбинированная оценка действий (например, «0-1-1») привела к трем различным классам ответного поведения: запоминание, передача и невосприимчивое поведение. Для построенных кривых мы использовали b 1 = 1, b 2 = 1, H e = 3,25 мВ и H i = 22 мВ.
https://doi.org/10.1371/journal.pone.0188003.g002
Оценка модели
В этом разделе мы описываем оценку предложенной канонической модели микросхемы в отношении (i) учета косвенной или прямой возбуждающей обратной связи, (ii) периодической обратной связи с IIN и (iii) баланса локальной сети, с помощью бифуркационных графиков и карт динамических функций.Мы показываем, что и при каких условиях модель поддерживает механизмы стробирования потока сигналов и рабочей памяти.
Основные динамики
Далее мы описываем ключевые особенности стробирующего механизма для трехпопуляционной модели, которая имеет косвенную возбуждающую обратную связь (рис. 1B). Эта конфигурация рассматривает отдельные нейронные массы для ввода и вывода. К популяции возбуждающих интернейронов (EIN) применялись прямоугольные всплески с различной интенсивностью и продолжительностью.Эти всплески имитировали входные данные из вышестоящих источников, таких как сенсорная информация, исходящая из первичных областей коры головного мозга, или информация более высокого уровня, такая как произнесенные слова или фонемы. Рис. 3A суммирует различные варианты поведения реакции: система реагирует на слабые и краткие стимулы небольшим отклонением мембранного потенциала Py (невосприимчивое поведение), но на более сильные, хотя и короткие, стимулы с большим переходным процессом, превышающим порог срабатывания 4 мВ. (поведение передачи). В обоих случаях система возвращается в исходное состояние вскоре после выключения стимула.Напротив, для более продолжительных стимулов система успокаивается в стабильном состоянии более высокой активации и остается нечувствительной к дальнейшим стимулам или шуму (поведение памяти). Находясь в этом селективном по стимулам высокоактивном состоянии, кратковременный ввод в IIN может активно сбросить систему в более низкое активированное состояние (S2 Рис.). Независимо от того, является ли ответ неотзывчивым, переносным или запоминающимся, зависит от выраженности применяемого стимула с точки зрения интенсивности и продолжительности, см. Рис. 3В. Диаграмма на рис. 3B служит характерным отпечатком пальца и отображает наблюдаемую динамику отклика.Соответствующие базовые операции стробирования и хранения могут служить строительными блоками для более сложных механизмов, таких как принятие решений, основанных на взаимодействии нейронов в единственной нейронной области. Полосатые узоры в переходной зоне между областями передачи и памяти на рис. 3В означают зависимость от времени выключения стимула относительно фазы собственных колебаний системы (см. S3 рис.).
Рис. 3. Аспекты реакции моделей на афферентные стимулы, поступающие на EIN трех популяционных моделей.
A) В зависимости от выраженности применяемых стимулов с точки зрения продолжительности и интенсивности наблюдались три различных поведения реакции: (1) невосприимчивое поведение после слабой и краткой стимуляции, когда реагирует мембранный потенциал Py, V Py . только с небольшим отклонением ниже порога срабатывания, см. импульс (i), (2) переходное поведение после сильной и краткой стимуляции, где V Py превышает порог срабатывания, см. импульс (ii) и (3) a поведение памяти после более длительной стимуляции средней интенсивности, см. импульс (iii), для которого система может установить стабильное состояние более высокой активации.В этом состоянии система нечувствительна к дальнейшим стимулам или шуму (см. Импульс (iv)), но может быть активно сброшена с помощью слабого и короткого импульса на IIN, очищая след памяти, см. Импульс (v). Обратите внимание, что этот импульс IIN был увеличен в 20 раз для улучшения четкости. Б) Реакция на поведение зависит от важности ввода. Невосприимчивое поведение показано для интенсивностей ниже 78 с -1 (зеленая область). Превышая эту интенсивность, более длинный стимул может надежно вызвать поведение памяти (оранжевая область).Чем короче стимул, тем более вероятно переходное поведение (серая область), где полосообразные узоры означают зависимость поведения (перенос или память) от фазового соотношения между временем выключения стимула и внутренними колебаниями системы. Для построенных кривых мы использовали b 1 = 1, b 2 = 1, H e = 3,25 мВ и H i = 22 мВ.
https://doi.org/10.1371/journal.pone.0188003.g003
Если посмотреть на базовую структуру пространства состояний, становится очевидным, что наблюдаемое поведение основано на довольно простом механизме.На рис. 4A визуализирована кривая с фиксированной точкой устойчивого состояния модели с тремя популяциями с параметризацией по умолчанию. Бифуркация складок была идентифицирована в каждой точке поворота кривой неподвижной точки — одной типа седло-узел (неустойчиво-стабильная) и одной типа седло-седло (нестабильная-нестабильная). Кроме того, подкритическая бифуркация Хопфа идентифицируется на p ext = -5.9s -1 . Получившаяся сепаратриса отмечает неустойчивое многообразие, которое отталкивает локальные траектории в пространстве состояний рядом с ним.Если в систему не подается входной сигнал (p ext = 0s -1 ), система находится на нижней ветви кривой с фиксированной точкой с V Py ≈ -2 мВ, см. Рис. 4A. Если к EIN приложен слабый импульс (p ext <78s -1 , см. Рис. 4B), система не сможет пройти бифуркацию нижней складки и снова установится на нижней ветви кривой фиксированной точки. . Если входной сигнал p ext > 78s -1 , система проходит бифуркацию нижней складки и устанавливается на верхней ветви кривой с фиксированной точкой, таким образом превышая порог срабатывания 4 мВ.Наличие пары отрицательных сопряженных комплексных собственных значений приводит к затухающим колебаниям. Когда стимул выключен, вход системы возвращается к исходному значению (p ext = 0s -1 ). Если траектория системы расположена за пределами сепаратрисы Хопфа (т.е. система не была достаточно демпфирована), система устанавливается на нижней ветви кривой фиксированной точки, реализуя переходное поведение (рис. 4C). Однако если траектория системы находится внутри сепаратрисы Хопфа (т.е. колебания достаточно затухают из-за достаточного времени установления), система останавливается на верхней ветви кривой с фиксированной точкой, демонстрируя, таким образом, поведение памяти (рис. 4D). Таким образом, отличительными признаками описанного механизма являются (i) бистабильная активация популяции Py (высокое и низкое состояние), (ii) селективность в отношении ярких стимулов, (iii) пониженная чувствительность к дальнейшим стимулам в высоком состоянии, (iv) относительная устойчивость к шумным колебаниям внешнего входа в каждом состоянии, и (v) фазовая зависимость смещения стимула, когда определенные фазы позволяют системе успокоиться в состоянии высокой активации, а другие фазы — нет (см. S3 Рис. Информация).
Рис. 4. Динамика различного поведения отклика в проекции пространства состояний.
A) S-образная кривая фиксированной точки показывает стабильные (сплошная линия) и нестабильную (пунктирная линия) фиксированные точки для различных входных значений EIN. Выявлены две бифуркации кратности (седло-узел и седло-седло) и докритическая бифуркация Хопфа. B-D) Проекции поведения отклика на бифуркационных диаграммах с входами, иллюстрирующими траектории в пространстве состояний и соответствующие временные ходы: неотзыв (B), переход (C) и поведение памяти (D).Обратите внимание, что V Py (t) = V 2 (t) -V 3 (t). Цветовое кодирование различает престимульный (красный), ответный (синий) и асимптотический (зеленый) средние мембранные потенциалы.
https://doi.org/10.1371/journal.pone.0188003.g004
В следующих подразделах мы оценим влияние структур положительной и отрицательной обратной связи на описанное поведение реакции. Для этого мы варьировали возбуждающие и тормозные синаптические усиления H e, i (контроль баланса сети).Для каждого варианта был сгенерирован характерный отпечаток пальца (см. Рис. 3B) и квалифицирован в пространстве состояний. Мы оценили три различные архитектуры в отношении баланса локальной сети: (i) архитектура с непрямой возбуждающей обратной связью без повторяющейся самопроверки IIN (рис. 1B), (ii) архитектура с прямой возбуждающей обратной связью без рекуррентной самопроверки IIN (рис. 1C). и (iii) архитектура прямой возбуждающей обратной связи с рекуррентной обратной связью IIN (рис. 1D).
Архитектура с косвенной возбуждающей обратной связью
Если оба параметра b 1 и b 2 установлены на единицу, контур возбуждающей обратной связи становится чисто косвенным (т.е.е., выходной сигнал Py подается в EIN, которые, в свою очередь, проецируются обратно на Py), и рекуррентная петля с обратной связью IIN исчезает (то есть трехпопуляционная модель, см. рис. 1B). На рис. 5 показана карта динамических функций, набор характерных отпечатков пальцев (рис. 3В). Карта динамической функции отображает классифицированное поведение отклика в пространстве параметров, охватываемом H e и H i , и показывает области, где в системе преобладают неотвечающие (ярко-зеленые, антрацитовые и голубые области), перенос (серые области) и поведение памяти (оранжевые и розовые области), а также их композиции.Соотношение возбуждения и торможения по умолчанию (H e = 3,25 мВ и H i = 22 мВ) [16], обозначающее баланс сети, расположено как раз на вершине большей области с преобладанием памяти (оранжевая область), соответствующей до уровней сильного торможения и близко к региону, где преобладает переносное поведение. Такая близость состояния системы к основным переходным зонам поведения системы (т. Е. Бифуркациям) была названа критичностью и считается благоприятной для способности системы обрабатывать информацию, поскольку небольшие изменения параметров могут привести к большим изменениям в поведении [47, 48 ].Следовательно, баланс локальной сети представляет собой очень чувствительный фактор, определяющий поведение канонической микросхемы, поскольку он настраивает критичность. Кроме того, баланс локальной сети контролирует перцептивную чувствительность системы в отношении интенсивности афферентных стимулов, которые воспринимаются или нет, путем настройки расстояния между рабочей точкой и бифуркацией нижней складки (сравните, например, S5G и S5H Fig). В параметризации по умолчанию (H e = 3,25 мВ и H i = 22 мВ) этот порог составлял примерно p ext = 78s -1 .Порог повышается при уменьшении H e , что означает снижение чувствительности к внешним раздражителям. В свою очередь, чувствительность увеличивается, когда этот порог понижается за счет увеличения H e . Обратите внимание, однако, что чистое увеличение H e приведет к поведению с преобладанием переноса, тогда как увеличение чувствительности в пользу поведения памяти требует одновременного уменьшения ингибирования (рис. 5).
Рис. 5. Карта динамических функций для архитектуры с непрямой возбуждающей обратной связью (см. Рис. 1B).Коллекция характерных отпечатков пальцев для различных возбуждающих (H e ) и тормозных (H i ) синаптических усилений. Цвета кодируют наблюдаемое поведение реакции: отсутствие реакции (ярко-зеленые, антрацитовые и голубые области), передача (серые области) и память (оранжевые и розовые области). Баланс локальной сети контролирует преобладание поведения и настраивает критичность системы. См. S4 Fig для дублирования этого рисунка, дополненного пояснительными диаграммами пространства состояний.
https://doi.org/10.1371/journal.pone.0188003.g005
Чтобы лучше понять механизм, лежащий в основе того, как баланс сети изменяет поведение ответа, мы дополнительно охарактеризовали систему в ее рабочей точке p ext = 0 Мы поддерживали внешний вход на нуле, систематически меняли H e и H i и отслеживали соответствующие бифуркации, как показано на рис. 6A. Фон графика окрашен в светло-красный цвет для колебательного поведения в низком состоянии при p ext = 0s -1 , светло-синий для не колебательного поведения и моностабильности и темно-синий для отсутствия колебаний и бистабильности.Значения параметров по умолчанию для H e и H i указаны на осях. Обратите внимание, что появляющиеся графики поразительно отражают границы областей поведения отклика с рис. 5. Синяя линия на рис. 6A указывает нижнюю бифуркацию складки для p ext = 0. Под этой линией эта разветвление будет располагаться по адресу p ext > 0 и выше на p ext <0 относительно p ext , сравните фиг. 6D и 6C. Точно так же голубая линия указывает верхнюю бифуркацию складки для p ext = 0s -1 , которая расположена на p ext <0 над этой линией и на p ext > 0 под линией, сравните Рис. 6D и 6G.Следовательно, только для области между двумя ветвями бифуркации складок точка p ext = 0s -1 будет расположена между двумя бифуркациями складок — необходимое условие для бистабильности в этой точке. Для поведения памяти необходимо, чтобы бистабильность существовала без ввода. Следовательно, две линии разветвления сгиба ограничивают область в плоскости H e -H i , где возможно поведение памяти. Это согласуется с наблюдаемым поведением, показанным на рис.5.Кроме того, между этими линиями расстояние между верхней и нижней разветвлениями складки на оси p ext (см. Рис. 6B – 6G) определяет устойчивость системы к шуму путем масштабирования ширины бистабильности. Более того, расположение системы относительно разветвления нижней складки определяет ее чувствительность к раздражителям. Однако не вся область между двумя ветвями бифуркации складок действительно проявляет бистабильность. Это связано с тем, что в какой-то момент, когда H i увеличивается, бифуркация верхней складки, переключение между нестабильными и стабильными фиксированными точками (бифуркация седло-узел, сплошная голубая линия на рис. 6A), разделяется на бифуркацию складки между двумя нестабильными фиксированными точками. точки (бифуркация седло-седло, пунктирная голубая линия) и докритическая бифуркация Хопфа, обозначенная пунктирной оранжевой кривой.Слева от этой кривой происходит докритическая бифуркация Хопфа при p ext <0, приводящая к бистабильности для p ext = 0 в виде устойчивого фокуса на верхней ветви кривой с фиксированной точкой, а справа от нее. Кривая бифуркации Хопфа находится при p ext > 0. Вследствие последнего верхняя ветвь кривой с фиксированной точкой нестабильна для p ext = 0, и бистабильность отменяется (рис. 6B). В какой-то момент вдоль этой ветви докритической бифуркации Хопфа докритическая бифуркация Хопфа превращается в суперкритическую бифуркацию Хопфа, обозначенную сплошной фиолетовой кривой на рис. 6A.Вместо стабильного фокуса устойчивый предельный цикл, связанный со сверхкритической бифуркацией Хопфа, затем учитывает бистабильность при p ext = 0. Вследствие этого существует также бистабильность вдоль узкой полосы на правой стороне сверхкритического значения Хопфа. бифуркация на рис. 6А, где верхнее состояние является колебательным.
Рис. 6. Двухпараметрический бифуркационный график трехпопуляционной модели.
A) График характеризует существующие бифуркации (относительно p ext ) в p ext = 0 для архитектуры непрямой возбуждающей обратной связи и отслеживает их через пространство параметров, охватываемое возбуждающим и тормозным синаптическим усилением H e и H i .Фон окрашен в светло-красный цвет для колебательного поведения в низком состоянии при p ext = 0s -1 , светло-синий для не колебательного поведения и моностабильности и темно-синий для отсутствия колебаний и бистабильности. Коричневые метки на оси обозначают значения параметров по умолчанию для H e и H i . Область между верхней (голубая линия) и нижней (синяя линия) бифуркациями складок и бифуркацией Хопфа (пурпурная / оранжевая кривая) демонстрирует бистабильность, где возможно поведение памяти, и H e и H i настраивают надежность и чувствительность система.Знаки + и – указывают, находится ли конкретная бифуркация складки в положительных или отрицательных значениях относительно рабочей точки. Сплошная фиолетовая линия обозначает закритическую ветвь бифуркации Хопфа. Пунктирная оранжевая линия обозначает докритическую ветвь бифуркации Хопфа, которая важна для переходного поведения системы. Вместе эти ветви обозначают границу между доминирующими областями памяти и поведения передачи (сравните S4 рис.). B) -G) Бифуркационные диаграммы, характеризующие стабильные и нестабильные фиксированные точки для широкого диапазона входных значений.
https://doi.org/10.1371/journal.pone.0188003.g006
Обратите внимание, что упомянутые докритические и сверхкритические бифуркации Хопфа сталкиваются при положительных / отрицательных значениях p ext в небольшой области, близкой к пунктирной / сплошной пурпурной линии соответственно (здесь не показаны). Это столкновение приводит к стабильному предельному циклу, который либо окружает небольшие биты перекрывающейся части кривой с фиксированной точкой (см. S5F рис.), Вызывая бистабильность и, следовательно, поведение памяти, либо достигает только нижней кратности, отменяя бистабильность и что приводит к поведению передачи (см. S5I рис.).В случае перекрытия, колебания этого глобального предельного цикла действительно пересекают порог срабатывания (классифицируется как роза на рис. S5F) или не пересекает порог срабатывания (обозначен оранжевым цветом на рис. S5G), и оба указывают на поведение памяти. Таким образом, темно-синяя область в плоскости H e -H i на рис. 6 демонстрирует бистабильность и допускает поведение памяти, но также является частью светло-синей области (из-за стабильных предельных циклов), что подтверждается рис. 5. Для настройки баланса сети за пределами этой бистабильной области каноническая микросхема больше не имеет поведения памяти и, таким образом, теряет неотъемлемую часть основных операций.
Архитектура с прямой возбуждающей обратной связью
Как объясняется в методике , мы можем легко транспонировать архитектуру непрямой возбуждающей обратной связи в архитектуру прямой возбуждающей обратной связи без каких-либо качественных изменений лежащей в основе корковой микроархитектуры. Для этой двухпопуляционной модели параметры равны b 1 = 0 и b 2 = 1 (см. Рис. 1C). Переход ограничен сохранением общего количества нейронов в системе, суммы протекающих токов и всех соединений между отдельными нейронами, как описано в файле S1.Двухпопуляционная модель не разделяет возбуждающие нейроны на входной и выходной слой. Для параметризации со значениями по умолчанию для H e и H i не наблюдалось бистабильности (см. Рис. 8A и 8E, в рабочей точке есть только одна стабильная фиксированная точка).
Карта динамических функций для архитектуры прямой возбуждающей обратной связи была получена и изображена на рис. 7. Поскольку колебательное поведение не наблюдается (в текущей рабочей точке p ext = 0, см. Рис. 8), эта карта демонстрирует меньше вариантов, так как по сравнению с архитектурой непрямой обратной связи (рис. 5), но все еще содержит все типы классифицированного поведения ответа, но для более низкого тормозящего синаптического усиления.Однако мы не смогли найти параметризацию, при которой все три типа поведения ответа устойчиво присутствуют в одном отпечатке пальца. Память и невосприимчивое поведение по-прежнему возникают в рамках одной параметризации и больше зависят от интенсивности, но не от продолжительности стимула. Кроме того, теряется способность реагировать затухающими колебаниями на дальнейший ввод, находясь в высоком состоянии. Теперь система сразу успокаивается. Как и в случае непрямой возбуждающей обратной связи, система в целом более чувствительна к изменениям возбуждающего синаптического усиления, чем к изменениям тормозного синаптического усиления.Это отражается в соответствующих диапазонах синаптических усилений, необходимых для получения желаемого поведения реакции.
Рис. 7. Карта динамических функций для архитектуры с прямой возбуждающей обратной связью (см. Рис. 1C).Коллекция характерных отпечатков пальцев для различных возбуждающих (H e ) и тормозных (H i ) синаптических усилений. Цвета кодируют наблюдаемое поведение реакции: отсутствие реакции (ярко-зеленый и антрацитовый), передача (серый) и память (оранжевый). Разнообразие наблюдаемого поведения сокращается по сравнению со случаем с тремя популяциями (рис. 5).Однако можно наблюдать все три основных типа. См. S5 Fig для дублирования этого рисунка, дополненного пояснительными диаграммами пространства состояний.
https://doi.org/10.1371/journal.pone.0188003.g007
Рис. 8. Двухпараметрический бифуркационный график модели с двумя популяциями.
График характеризует существующие бифуркации в p ext = 0s -1 для архитектуры прямой возбуждающей обратной связи и отслеживает их через пространство параметров, охватываемое возбуждающими и тормозными синаптическими усилениями H e и H i .Область между верхней (голубая линия) и нижней (синяя линия) ветвью бифуркации сгиба ограничивает область, где получается бистабильная кривая с фиксированной точкой. Однако докритическая бифуркация Хопфа (фиолетовая линия) делает части кривой с фиксированной точкой нестабильными и предотвращает фактическую бистабильность при p ext = 0s -1 , что подавляет поведение памяти в пользу поведения передачи (см. Рис. ).
https://doi.org/10.1371/journal.pone.0188003.g008
Что касается архитектуры косвенной возбуждающей обратной связи, существование бифуркаций было исследовано в рабочей точке p ext = 0s -1 и изображено на Фиг.8А.Опять же, ветви бифуркации отражают границы областей поведения реакции в расширенной карте динамических функций с рис. 7. Как описано для архитектуры косвенной обратной связи, поведение бистабильности и памяти обычно существует в области между ветвями бифуркации верхней и нижней складок, пока они отражают бифуркации седло-узел. Однако, также как и в случае косвенной обратной связи, примерно при H — = 6 мВ верхняя бифуркация складки (голубая кривая на рис. 8A) разделяется на седло-седло и докритическую бифуркацию Хопфа, и только слева от этой бифуркации Хопфа. бистабильность существует, сравните рис. 8C и 8D.В общем, мембранные потенциалы в высоком состоянии довольно высоки (> 10 мВ) и близки к порогу насыщения сигмовидной функции около 14,8 мВ. Эфферентные скорости возбуждения таких высоких мембранных потенциалов, следовательно, будут приводить к почти неизменно передаваемым скоростям возбуждения 5 с -1 . Как следствие, ступенчатые эфферентные скорости возбуждения, как и в случае архитектуры непрямой возбуждающей обратной связи, являются довольно исключительными.
Таким образом, двухпопуляционная модель демонстрирует все типы ответного поведения, но для более низких диапазонов тормозящего синаптического усиления.Одновременного существования всех типов поведения ответа для одного значения баланса локальной сети не наблюдалось.
Архитектура прямой возбуждающей обратной связи с растормаживанием
Часто игнорируемым свойством нейронных архитектур является наличие повторяющихся тормозных коллатералей, приводящих к тормозящей самооценке и приводящих к эффекту растормаживания. В то время как ингибирование ослабляет возбужденную систему, растормаживание уменьшает это демпфирование и снижает эффективность тормозного пути обратной связи.Это способствует насыщению возбуждающего тракта обратной связи и может привести к стационарной динамике, при которой мембранный потенциал Py может возрастать в линейной зависимости от внешнего входа. Этот эффект еще более усиливается при высоком возбуждающем (усиление положительной обратной связи) и тормозящем (усиление растормаживания) синаптических усилиях.
Мы ввели растормаживание в двухпопуляционную модель, установив архитектурный параметр b 2 на ноль, сохранив b 1 = 0 (см. Рис. 1D), медленно увеличили растормаживание, контролируя усиление связности N II , и отследили существующие бифуркации. в пространстве состояний для двухпопуляционной модели (см. S1 рис.).Поскольку динамика существенно не изменилась для больших уровней растормаживания, мы зафиксировали N II и N II = 33,25, что соответствует уровню ингибирующих соединений, нацеленных на возбуждающие популяции.
Карта динамических функций для архитектуры прямой возбуждающей обратной связи с тормозящей рекуррентной обратной связью была получена и показана на рис. 7. Опять же, наблюдались все типы классифицированного ответного поведения. Для некоторых параметризаций наблюдались два поведения в зависимости от интенсивности стимула.Опять же, система была более чувствительна с точки зрения различного поведения реакции к изменениям возбуждающего синаптического усиления, чем к изменениям тормозного синаптического усиления. Как и раньше, бифуркации в рабочей точке p ext = 0 в пространстве H e -H i , которые отражают границы областей ответного поведения на карте динамических функций для двухпопуляционной модели с растормаживанием. (Рис. 7), были исследованы и изображены на Рис. 8. Он показывает, что для увеличения тормозящего синаптического усиления бифуркации верхней и нижней складок, которые ограничивают диапазон мультистабильности кривой с фиксированной точкой, больше напоминают трехпопуляционную модель, чем две -популяционная модель без растормаживания.Более того, бифуркация верхней складки остается бифуркацией седлового узла для увеличения тормозных синаптических усилений и сохраняет стабильность верхней части кривой фиксированной точки. Это контрастирует с обеими альтернативными моделями, для которых бифуркация седло-узел расщепляется на седло-седло и докритическую бифуркацию Хопфа (рис. 8). Сохранившаяся бистабильность и отсутствие сепаратрисы (отфильтровывающая краткие стимулы, см. Рис. 6) объясняют значительно больший диапазон рабочей памяти в двухпопуляционной модели с растормаживанием.Что касается стробирования потока сигнала, модель с двумя популяциями с растормаживанием фильтрует стимулы в соответствии с их интенсивностью, но игнорирует их временную согласованность и быстротечность.
Рис. 9. Карта динамической функции для двухпопуляционной модели с растормаживанием (см. Рис. 1D).Коллекция характерных отпечатков пальцев для различных возбуждающих (H e ) и тормозных (H i ) синаптических усилений. Наблюдаемое поведение реакции обозначено цветом: отсутствие реакции (ярко-зеленый и антрацитовый), перенос (серый) и память (оранжевый).Разнообразие наблюдаемого поведения сокращается по сравнению со случаем с тремя популяциями (рис. 5). Однако можно наблюдать все три основных типа. См. S6 Fig для дублирования этого рисунка, дополненного пояснительными диаграммами пространства состояний.
https://doi.org/10.1371/journal.pone.0188003.g009
Рис. 10. Двухпараметрический бифуркационный график для двухпопуляционной модели с растормаживанием (см. рис. 1D).A) График характеризует существующие бифуркации в p ext = 0 для архитектуры прямой возбуждающей обратной связи с растормаживанием и отслеживает их через пространство параметров, охватываемое возбуждающими и тормозными синаптическими усилениями H e и H i .Область между верхней (голубая линия) и нижней (синяя линия) бифуркацией складки ограничивает диапазон параметров для бистабильной кривой с фиксированной точкой. Эти диапазоны ветвей бифуркации отражают границы поведения невосприимчивости, передачи и памяти на рис. 7). B) -D) Однопараметрические бифуркационные графики показывают кривую с фиксированной точкой (V Py ) и локальные бифуркации вдоль p ext для различных значений баланса локальной сети.
https://doi.org/10.1371/journal.pone.0188003.g010
Приложение для обработки предложений
Мы использовали каноническую микросхему для моделирования когнитивной функции обработки предложений. Во время восприятия предложения непрерывный поток слов постепенно преобразуется в иерархически организованное нейронное представление, отражающее значение воспринимаемого предложения. Воспроизведение предложения через некоторое время требует локального представления, в котором хранятся слова и их отношение друг к другу.В этой части исследования мы показываем, как сеть, состоящая из канонических микросхем, может анализировать предложение на основе его синтаксической информации. Выборочная активация нейронных областей, представляющих слова, и определенное сохранение локальной информации основываются на основных операциях, рассмотренных выше. Далее мы показываем, как изменение баланса сети нарушает процесс построения структуры восприятия предложения.
Ниже мы рассмотрим вопрос, как неоднозначная словесная информация в примерном предложении « Я ударил вора дубиной » может быть представлена в распределенной нейронной сети канонических микросхем.Это предложение неоднозначно по синтаксису: фразу « с дубиной » можно интерпретировать как наречие, то есть дополнительно определяющее « хит» , или как прилагательное, дополнительно определяющее « вор» . Мы предполагаем, что эта неоднозначность разрешается доступной контекстной информацией, то есть просодической информацией или конкретными знаниями, касающимися дискурса или темы. В предлагаемой сетевой модели, изображенной на рис.9, каждое слово представлено одной нейронной областью (т.е., позиционное кодирование см. [49]), смоделированное через единую каноническую микросхему. Эти микросхемы, представляющие слова, подразделяются на модули в соответствии с их синтаксической ролью, то есть субъектами, глаголами, объектами и их модификаторами. Значения внешних межмодульных и внутримодульных соединений были оптимизированы вручную, чтобы дать разумные ответы. Эти значения можно найти на рис. 10. Далее, мы предполагаем, что временной порядок слов предоставляет информацию о назначении субъекта и объекта [49].Предлагаемый расчет построения структуры основан на управляемой вводом последовательной активации канонических микросхем. Активированная микросхема, принадлежащая определенному словарному модулю, передает свою увеличенную частоту срабатывания тем модулям, которые, вероятно, последуют, и по-разному предварительно активирует соответствующие слова посредством взвешенных соединений, создавая ожидания. В модели эта ступенчатая предварительная активация соответствует сдвигу в базовой активации микросхемы и приближает систему к соответствующей складчатой бифуркации (см. Раздел «Основная динамика», и рис. 4).В этом смысле активация слова « съесть » в модуле глаголов предварительно активирует слова в модуле глаголов-модификаторов и в модуле объектов, но не активирует снова слова в модуле глаголов ( см. рис.9). Информация о последующих афферентных словах может затем полностью активировать соответствующую микросхему и продолжить процесс построения структуры. Модули модификации глагола и объекта (см. Рис. 9) допускают конкурирующие интерпретации предложения. Их взаимное подавление в сочетании с текущим уровнем контекстной информации гарантирует, что единовременно поддерживается одна конкретная интерпретация.Топология сети включает данные о фразовой структуре предложений, принципах синтаксического анализа (например, позднее закрытие, [50]) и отражает предсказательный характер обработки предложений.
Рис. 11. Сеть обработки предложений для понимания предложений.
Информация об афферентном слове выборочно возбуждает каноническую микросхему, представляющую слово, когда соответствующее слово распознается в первичных слуховых областях. Активированная микросхема, например, представляющая слово ‘ I ‘ и принадлежащая предметному модулю (S), предварительно активирует слова в подключенном глагольном модуле (V) и вместе с выборочной информацией афферентного слова активирует другую микросхему. (‘ есть ‘).Теперь слова как в модуле глаголов-модификаторов (V мод.), Так и в модуле объектов (O) дифференцированно предварительно активируются взвешенными связями. Контекстная информация предлагается для управления этим управляемым вводом процессом построения структуры путем модуляции возбудимости целевой микросхемы, например, в нашем случае, посредством ингибирования.
https://doi.org/10.1371/journal.pone.0188003.g011
Рис. 12. Нейронное представление воспринимаемого предложения распределенной сетью из шести взаимодействующих канонических микросхем.
A) Топология сети включает модули (пунктирные прямоугольники), содержащие шесть соответствующих канонических микросхем, представляющих слова (сплошные прямоугольники), которые связаны между собой посредством возбуждающих и тормозящих связей индивидуальной силы (B). Цвета линий последовательно отражают соответствующие слова на всех панелях. C) Интерпретация, в которой фраза « with the club » относится к фразе прилагательного, руководствуется контекстной информацией (например, знанием того, что есть вор с дубинкой), которая препятствует модулю модификаторов глаголов.D) Для второй интерпретации, интерпретации « с клубом » как наречия, контекстная информация невелика, и модуль изменения глагола остается активным. E, F) В случае, если баланс локальной сети микросхем смещен в пользу тормозящих влияний, точное построение структуры не удастся для обеих интерпретаций (E и F) и приведет к неправильной интерпретации, то есть к дефектным следам активации слов (верхние графики), или потеря памяти, т.е. отсутствие длительного следа активации (нижние графики).
https://doi.org/10.1371/journal.pone.0188003.g012
Для представления примерного предложения « Я ударил вора дубинкой » мы сосредоточились на соответствующих словах и игнорировании связи с не задействованными микросхемами. . Следовательно, мы создали сеть из шести взаимодействующих канонических микросхем (см. Рис. 10A), каждая из которых смоделирована трехпопуляционной моделью (рис. 1B), и их соответствующих соединений, см. Рис. 10B. Параметры см. В таблице 1. Неопределенность, т.е.е. вопрос о том, служит ли фраза « with the club » прилагательным или наречием, решается путем отделения модулей модификации глагола от модулей модификации объекта. Оба модуля взаимно подавляют друг друга через асимметричные соединения, см. Рис. 10B. Кроме того, текущий уровень контекстной информации направляет процесс построения структуры. При моделировании контекстная информация моделировалась как тормозящий шумовой сигнал с постоянным смещением, которое ухудшает афферентный вход, представленный p ext (см. Рис. 1), целевой области.Наблюдались две интерпретации одного и того же афферентного слова. Для первой интерпретации (см. Фиг. 10C) словесная информация активирует « I », а затем « hit ». Это предварительно активирует как объектный модуль, так и модуль глагола-модификатора. Информация по афферентному слову теперь активирует « the thief », который предварительно активирует модуль объектного модификатора. Информация афферентного слова затем активирует « с » как в модуле изменения глагола, так и в модуле изменения объекта, что приводит к конкурентному взаимодействию, которое устраняет двусмысленность.Во время соревнований контекстная информация блокирует модуль изменения глаголов. Дополнительная информация афферентного слова активирует « клуб» и завершает процесс построения структуры. Для второй интерпретации (см. Рис. 10D) конкретная контекстная информация отсутствует, так что интерпретация происходит в соответствии с опытом слушателя, то есть индивидуальным соотношением взаимного торможения между модификаторами. Следовательно, модификация глагола запрещает модуль модификации объекта, и « с дубиной » интерпретируется как наречивая фраза.
Некоторые неврологические расстройства связаны с нарушением баланса сети на уровне взаимодействующих нейронных популяций [26, 29, 32]. Кроме того, известно, что лекарства, анестетики и другие химические вещества изменяют количество или эффективность доступных рецепторов нейромедиаторов. Валенсуэла [51] сообщает о нарушении баланса сети в пользу тормозящих влияний после употребления алкоголя. В модели с тремя популяциями мы воспроизвели этот сценарий, слегка увеличив тормозной синаптический прирост с H i = 22 мВ до H i = 23 мВ (H i = 24 мВ) и наблюдали дефектное представление предложения (см. Рис. 10E и 10F).Хотя представляющие канонические микросхемы получают информацию о соответствующем афферентном слове, они не могут проявлять необходимую устойчивую активность.
Таким образом, сеть взаимодействующих канонических микросхем способна преобразовывать поток информации афферентного слова в репрезентативный след нейронной активности посредством управляемой вводом последовательной активации. Однако, если канонические микросхемы не могут выполнять необходимые базовые операции, например, из-за расстройки баланса сети, этот процесс построения структуры завершится неудачей.
Обсуждение
В этом исследовании мы демонстрируем, как локальная архитектура коры, представленная канонической микросхемой, реализует основные операции обработки информации стробирования потока сигналов и рабочей памяти. Эти базовые операции формируют основные предпосылки для корковых вычислений более высокого порядка, таких как построение структуры. Мы исследовали, в какой степени локальный топологический выбор ограничивает эти основные операции. Мы продемонстрировали, что только модели с отдельными возбуждающими входными и выходными популяциями (т.е., трехпопуляционная модель с косвенной возбуждающей обратной связью) одновременно учитывают все соответствующие варианты поведения для единой параметризации. Важно отметить, что баланс локальной сети оказался решающим фактором для доступности основных операций и, следовательно, функциональной индивидуализации микросхемы. Мы продемонстрировали, как сеть, состоящая из нескольких индивидуальных канонических микросхем, может реализовать сложную когнитивную операцию, а именно синтаксический анализ при обработке предложений.
Далее мы обсудим, как наш выбор основных операций и канонической модели микросхемы может рассматриваться как общий знаменатель для различных других утверждений, встречающихся в литературе. Мы будем утверждать, что модели нейронной массы являются подходящим выбором для представления канонической микросхемы, и укажем, как наши исследования выходят за рамки предыдущих исследований моделей нейронной массы. Более того, мы разъясним сильные стороны и ограничения представленного примера в свете нейролингвистики, а также других вычислительных моделей обработки предложений.
Основные операции в канонических микросхемах
Представление о разнообразных когнитивных функциях, основанных на относительно однородной локальной архитектуре нижележащего нейронного субстрата, поддерживает идею канонических микросхем [6, 52], эффективное взаимодействие которых составляет вычислительную мощность коры головного мозга [1–3]. На данный момент нейробиологические исследования согласуются с такими характеристиками коры головного мозга, как ламинация, биофизические свойства доминирующих типов нейронов, а также целевые и исходные слои передаваемых сигналов [6, 53].Заметим, однако, что эта концепция не совсем бесспорна [54, 55].
Канонические микросхемы ранее ассоциировались с базовыми операциями или стереотипными функциями [9], такими как управление усилением и восстановление сигнала [56], линейными (например, суммирование, деление и инверсия знака) и нелинейными операциями (например, победитель получает- все, инвариантность и мультистабильность) [7], усиление и нормализация сигнала [55, 57], а также селективность и вычисление коэффициента усиления [2]. Крупномасштабные импульсные нейронные сети, явно имитирующие ламинарную архитектуру кортикального столба, широко используются для изучения связи между стереотипной структурой и базовыми операциями в вычислительных моделях канонических микросхем [8, 58–60].Наличие таких операций также исследовалось в моделях среднего поля [61–63], которые составляют основу большинства попыток моделирования нейрокогнитивных экспериментов (например, DCM; [37, 64]). В настоящем исследовании мы продвигаем понимание канонических операций, предоставляя доказательства фундаментальных базовых операций даже в очень простой модели обратной связи канонической микросхемы. Из-за своего фундаментального характера идентифицированные базовые операции (стробирование потока сигналов и рабочая память) подразумевают другие ранее описанные канонические операции (см. Выше), которые являются более сложными и были обнаружены в очень специфических архитектурах кортикального столбца.Установлено, что модели среднего поля могут надежно описывать коллективное поведение большого числа нейронов [65]. Важно отметить, что пространственная абстрактность выбранного нами типа модели, гибко отражающей взаимодействие нескольких нейронов, нейронных популяций или целых областей коры, позволяет интерпретировать эту базовую функциональность, которая актуальна на различных уровнях нейронной организации, потенциально превышая уровень коркового столба.
Тем не менее, мезоскопический пространственный масштаб нейронных популяций, в котором, возможно, устанавливается однородность канонических микросхем, хорошо охвачен концепцией моделей нейронной массы [20, 65].Здесь мы исследуем его динамику в ответ на переходный вход в свете изменений основного пространства состояний, что наиболее важно для обработки информации, управляемой стимулами.
Наше внимание к отдельной микросхеме дополняет недавние разработки в области сетевых технологий, которые также обращаются к идее моделирования канонических микросхем с помощью моделей нейронных масс. Было изучено, как очевидное различие межсхемных соединений на прямые и обратные связи, с ламинарно-специфическим происхождением и целями [22, 38, 66], мотивирует расположение канонических микросхем в иерархических моделях [37, 67, 68] .Что еще не изучено в этих моделях, так это основные операции на уровне микросхемы и их чувствительность к топологическим особенностям и уровням возбуждения и торможения. В этом исследовании мы исследовали поведение реакции, вызванной стимуляцией, в трех репрезентативных локальных топологиях: (i) трехпопуляционная модель (рис. 1B), (ii) двухпопуляционная модель (рис. 1C) и (iii) двухпопуляционная модель (рис. 1C) и (iii) двухпопуляционная модель. популяционная модель с повторяющейся тормозной обратной связью (рис. 1D). Среди рассмотренных архитектур обратной связи и соответствующих диапазонов параметров мы обнаружили, что только модель с тремя популяциями (с входными данными для возбуждающих интернейронов, таким образом, с отдельными входными и выходными популяциями) демонстрирует сосуществование всех трех вариантов поведения ответа для фиксированного значения баланса сети. .То есть только трехпопуляционная архитектура способна выборочно блокировать, передавать или запоминать стимул в зависимости от его свойств. Кроме того, поведение переноса моделей с двумя популяциями зависит только от силы применяемых стимулов, а не от их продолжительности, в отличие от модели с тремя популяциями, где оно зависит от обоих. В этом смысле наши результаты демонстрируют, что рассмотрение косвенной возбуждающей петли обратной связи очень важным образом увеличивает разнообразие и биологический реализм динамики модели.Поскольку более сложные архитектуры моделей обычно демонстрируют более богатый динамический репертуар, мы ожидаем, что такое поведение также будет иметь место в более детальных моделях канонических микросхем. В частности, наша модель не учитывает прямое взаимодействие между возбуждающими и тормозными интернейронами, а также саморегуляторную связь между возбуждающими интернейронами. Обе эти связи, безусловно, существуют [58]. Эти особенности являются одними из важных расширений модели, которые необходимо изучить в будущих исследованиях.
В нашей модели канонической микросхемы мы описали поведение локальной памяти на основе бистабильности. Повторяющаяся и самоподдерживающаяся активность модулируется балансом сети и инициируется и прекращается афферентными входами, поведение, которое было показано in vitro [35]. В отличие от долговременной памяти, которая опирается на синаптическую пластичность, краткосрочная (или рабочая) память, как полагают, полагается на механизмы, которые не изменяют основную структуру связности. В качестве альтернативы было предложено, что рабочая память может быть основана на задержках и постоянных времени [12, 69], таких как цепочки синфайров , [70], повторяющиеся возбуждающие сети [71] или свойства клеток [72].В этих механизмах период хранения (время забывания) зависит от относительно фиксированных структурных аспектов сети, тогда как при бистабильности элемент может храниться в памяти в принципе в течение любого периода, пока он не будет активно выключен (например, с помощью импульса ингибирующая популяция, см. S2 фиг.).
Нейронные функции должны быть устойчивы к шуму, но в то же время чувствительны к афферентному входу. Сигналы можно отличить от шума по более высокой амплитуде и плавности во времени.В нашей модели по амплитуде расстояние между рабочей точкой и бифуркацией складки должно быть больше шума и меньше сигнала. Мы продемонстрировали, что это расстояние эффективно контролируется балансом локальной сети, который, следовательно, является ключевым параметром для управления компромиссом между устойчивостью к шуму и чувствительностью к стимулам (например, S4 Fig). Что касается временной гладкости, только трехпопуляционная модель предлагает механизм отбора. Мы установили, что модель сохраняет стимул в памяти только в том случае, если он длится достаточно долго, то есть демонстрирует большую степень временной плавности.
В дополнение к концепции функциональной диверсификации по топологии [3], мы показали, что изменение баланса локальной сети также является биологически приемлемым средством индивидуализации локальной функциональности. Эта локальная настройка определяет, будет ли нейронная область выборочно пересылать импульс (поведение передачи), переключаться на более высокую постоянную активацию (поведение памяти) или вообще не реагировать на импульс (невосприимчивое поведение). Отметим, что в этом случае мы рассматривали баланс локальной сети как сосредоточенный параметр, который фактически инкапсулирует большое количество физиологических и анатомических свойств и механизмов, таких как кинетика нейротрансмиттеров и плотности нейрорецепторов, дендритное ветвление, синаптическая и внеклеточная ионная динамика и подключения к локальной и нелокальной сети, и это лишь некоторые из них [73, 74].Из-за этой двусмысленности и, как обсуждалось ранее, концепцию баланса сети очень трудно дать количественной оценке. В этом исследовании мы использовали простое определение, чтобы продемонстрировать в качестве доказательства концепции, что баланс локальной сети действительно обеспечивает фундаментальное средство индивидуализации функциональности локальной микросхемы.
Применение канонической микросхемы для обработки предложений
Идея обширной сети подобных микросхем, поддерживающих функцию коры головного мозга, была выдвинута в специализированных нейрокогнитивных теориях.Например, было выдвинуто несколько предложений, чтобы обеспечить механистическое понимание обработки языка [1, 49, 61, 62, 75]. Это также было связано с организационными принципами, такими как кодирование места, где нейронная область представляет собой абстрактный элемент обработки [49]. В этом исследовании мы использовали канонические микросхемы для представления составляющих воспринимаемых предложений, отражая принцип кодирования мест. Канонические микросхемы, представляющие одно слово, были сгруппированы в модули в соответствии с их синтаксической ролью (например,g., субъект, объект), а не их словесную категорию (например, существительное, местоимение). Мы описываем вычислительный механизм управляемой вводом функциональной привязки дискретных элементов. Это позволяет генерировать бесконечные последовательности из ограниченного числа дискретных элементов, концепция, называемая бесконечной рекурсией [3]. Предлагаемый механизм построения генеративной структуры можно было бы назвать динамическим набором , чтобы подчеркнуть свободу структуры в отношении ее количества и порядка элементов.
Предлагаемый вычислительный механизм применяется для синтаксического анализа, то есть группирования непрерывного потока слов в иерархическую структуру составных частей предложения. Мы ограничиваем механизм построения структуры, фиксируя лежащую в основе проводку взаимодействующих нейронных областей. Характеристики сети, то есть ее топология и веса соединений, отражают правила синтаксиса, которые, вероятно, будут установлены во время изучения языка. Базовые операции, обеспечиваемые канонической микросхемой, имеют решающее значение для этого механизма построения структуры.Очевидно, что для реализации временной интеграции последовательностей в реальном времени необходим эффективный механизм краткосрочной памяти. Механизм рабочей памяти нашей канонической микросхемы обеспечивает быстрое кодирование и гибкое время хранения. Последнее особенно актуально для учета разной скорости речи и разной длины предложений. Стробирование потока сигналов гарантирует, что слова активируются только в том случае, если они были предварительно активированы (ожидаемы) структурой предложения и распознаны из входного потока.Поскольку было показано, что эти базовые операции в значительной степени зависят от баланса сети, изменение этого параметра приводит к сбою работы глобальной сети. В текущей реализации языковой сети сосуществование памяти и поведения передачи, которое является отличительной чертой модели трех популяций, не является необходимым ингредиентом. Таким образом, также могла быть использована модель двух популяций. Однако для дальнейшей разработки модели могут стать актуальными специфические свойства модели трех популяций.Например, из-за неспецифической информации афферентного слова отдельные слова в принципе могут встречаться несколько раз (например, слово пить как субъект, глагол, объект). Чтобы предотвратить одновременную устойчивую активацию нескольких канонических микросхем (которая приведет к насыщению узлов и сделает их недоступными для последующей активации), локальная схема должна иметь поведение передачи, обеспечиваемое моделью трех популяций.
Предлагается, чтобы в процессе понимания предложения за первоначальным синтаксическим анализом, рассматриваемым в данном исследовании, следовало тематическое распределение ролей и коллективная оценка синтаксической и семантической информации [76].Помимо прагматической информации, предлагается также просодическая информация для руководства пониманием предложения [77]. В нашей модели мы объединяем все типы такой дополнительной информации в более общее понятие контекстной информации.
Предлагаемая сетевая модель отражает характеристики модели serial , syntax-first путем построения простейшей синтаксической структуры на основе информации категории слова, независимой от лексико-семантической информации [76]. Модель также отражает характеристики моделей удовлетворения ограничений [78] за счет включения неструктурных факторов в топологию сети.К таким факторам относятся частота конкретной структуры или ее семантическая правдоподобность. В соответствии с концепцией моделей удовлетворения ограничений, предлагаемая сетевая модель способна учитывать синтаксические неоднозначности (см. Рис. 10). Мы также предполагаем, что восприятие и идентификация словесной информации информирует, но концептуально отличается от вычислений построения структуры. Входящая слуховая информация распознается как слово [79], которое затем вводится в нейронные области, которые задействуются во время синтаксического анализа.
В дополнение к аналогичной модели [49] наша модель рассматривает модификаторы глаголов и объектов. Однако в предложенном здесь механизме построение структуры все еще сильно опирается на временные порядки слов, то есть субъект-глагол-объект. Помимо информации о категории слов, успешный синтаксический анализ также должен учитывать морфосинтаксические особенности, такие как пол, префиксы или падежи. Каким образом эти функции встроены — вопрос открытый и требует дальнейшего изучения.
Одной из целей нашего исследования было проложить путь к механистическому пониманию обработки предложений в соответствии с нейропсихологическими теориями.Это противоположно области компьютерной лингвистики, которая направлена на оптимизацию разговорного взаимодействия человека с машиной, жертвуя при этом биологической достоверностью. Кроме того, мы хотели бы подчеркнуть характер развития нашей модели и осознаем несколько недостатков, некоторые из которых мы устраняем здесь:
На уровне слов.
Трудно справиться с многократным упоминанием одного и того же слова в предложении. Как только слово в отдельном модуле активируется, его необходимо деактивировать, прежде чем его снова можно будет активировать.Хотя механизм деактивации неявно включен в нашу модель, через короткий импульс к IIN, деактивация в режиме онлайн одного слова прервала бы репрезентативную трассировку активации. Кроме того, суммирующий эффект нескольких предварительных активаций в конечном итоге приведет к активации области, даже если слово отсутствует на входе. Расширение модели могло бы масштабировать уровень предварительной активации относительно порога активации, так что слово «на кончике языка», но еще не активировано.
На уровне приговора.
Опять же, представления необходимо деактивировать, прежде чем можно будет представить следующее предложение. Чтобы следить за беседой, необходимо активировать последовательность слов и связанные с ними словесные сети [80]. Эти сети медленно распадаются, так что информация, охватывающая несколько предложений, может быть связана вместе посредством ассоциативных процессов. Наконец, ранее было показано, что сети со слишком большим количеством активированных узлов имеют тенденцию становиться нестабильными и тем самым уничтожать информацию, хранящуюся в состоянии сети [42], которая может иметь отношение к длинным или сложным предложениям.
Заключение
Наши результаты подтверждают концепцию вычислительных примитивов [7] или стереотипных функций [9] и определяют минимальную структуру модели для канонической микросхемы, которая поддерживает эти функции. Они также подтверждают на локальном уровне решающую роль баланса сети для способности нейронных сетей обрабатывать информацию. Мы пришли к выводу, что наши результаты подтверждают коннекционистскую идею о том, что более высокая функция мозга возникает из сетей относительно похожих, хотя и настраиваемых индивидуально, канонических микросхем.
Вспомогательная информация
S1 Рис. Увеличение повторяющейся тормозящей самовозвратной связи в двухпопуляционной модели.
A) Двухпараметрический график бифуркации отслеживает возникающие бифуркации вдоль p ext , когда увеличивается повторяющаяся тормозящая самовозвращающая обратная связь N II (то есть, b 2 установлено на ноль, см. Рис. 1D). Баланс сети поддерживался постоянным при значениях H e = 3,25 мВ и H i = 22 мВ. B-E) Однопараметрический график бифуркации показывает кривую с фиксированной точкой (V Py ) и локальные бифуркации вдоль p ext для различных значений N II .F) Кривые с фиксированной точкой для скорости срабатывания Py, φ (t), вдоль p ext для различных значений баланса локальной сети.
https://doi.org/10.1371/journal.pone.0188003.s001
(TIF)
S3 Рис. Фазовая зависимость между собственными колебаниями системы и временем выключения стимула.
На диаграмме показан набор ответов системы на стимулы постоянной интенсивности (100 с -1 ) и длительностью стимула в диапазоне 600-750 мс, где время смещения стимула отмечено вертикальными линиями.Синие линии обозначают ответы на стимулы, для которых система в конечном итоге вернется в неактивное состояние после выключения стимула (то есть поведение передачи). Красные линии обозначают стимулы и ответы, при которых система могла оставаться активированной (то есть поведение памяти). Останется ли система в активированном состоянии, зависит от момента времени смещения стимула относительно фазы колебательного ответа. Такое поведение возникает из-за четкой траектории системы в пространстве состояний при включенном стимуле.Как только стимул выключен, фазовая точка системы либо находится в области притяжения устойчивого фокуса верхней ветви кривой с фиксированной точкой, либо будет притягиваться к устойчивому узлу нижней ветви кривой с фиксированной точкой. (сравните с рис. 4). Обе области притяжения разделены сепаратрисой неправильной формы, возникающей в результате нестабильной бифуркации Хопфа (см. Проекцию на рис. 4A). Чем больше длительность стимула, тем больше времени у системы есть на то, чтобы успокоиться до кривой фиксированной точки, что увеличивает вероятность пребывания в зоне притяжения фиксированной точки верхней ветви (поведение памяти) и вызывает более широкую полосы для большей продолжительности стимула на рис. 3B.
https://doi.org/10.1371/journal.pone.0188003.s003
(TIF)
S4 Рис. Карта динамических функций для архитектуры с непрямой возбуждающей обратной связью (см. Рис. 1B).
A) Набор характерных отпечатков пальцев для различных возбуждающих (H e ) и тормозных (H i ) синаптических усилений. Цвета кодируют наблюдаемое поведение реакции: отсутствие реакции (ярко-зеленые, антрацитовые и голубые области), передача (серые области) и память (оранжевые и розовые области).Баланс локальной сети контролирует преобладание поведения и настраивает критичность системы. BJ) Примерные параметризации, показывающие отпечатки пальцев, временные ходы и их проекции на бифуркационном графике.
https://doi.org/10.1371/journal.pone.0188003.s004
(TIF)
S5 Рис. Карта динамических функций для архитектуры с прямой возбуждающей обратной связью (см. Рис. 1C).
A) Набор характерных отпечатков пальцев для различных возбуждающих (H e ) и тормозных (H i ) синаптических усилений.Цвета кодируют наблюдаемое поведение реакции: отсутствие реакции (ярко-зеленый и антрацитовый), передача (серый) и память (оранжевый). Разнообразие наблюдаемого поведения сокращается по сравнению со случаем с тремя популяциями (S4, рис.). Однако можно наблюдать все три основных типа. B) -G) Выбранные параметризации, показывающие отпечатки пальцев, временные ходы и их проекции на бифуркационном графике.
https://doi.org/10.1371/journal.pone.0188003.s005
(TIF)
S6 Рис. Динамическая функциональная карта для двухпопуляционной модели с растормаживанием (см. Рис. 1D).
A) Набор характерных отпечатков пальцев для различных возбуждающих (H e ) и тормозных (H i ) синаптических усилений. Наблюдаемое поведение реакции обозначено цветом: отсутствие реакции (ярко-зеленый и антрацитовый), перенос (серый) и память (оранжевый). Разнообразие наблюдаемого поведения сокращается по сравнению со случаем с тремя популяциями (рис. 5). Однако можно наблюдать все три основных типа. B) -E) Выбранные параметризации с отпечатками пальцев, временными курсами и их проекциями на бифуркационном графике.
https://doi.org/10.1371/journal.pone.0188003.s006
(TIF)
Благодарности
Мы благодарим Йенса Брауэра за полезные комментарии к рукописи.
Ссылки
- 1. Friederici AD, Singer W. Основание языковой обработки на основных нейрофизиологических принципах. Тенденции в когнитивных науках. 2015; 19 (6): 329–38. Epub 2015/04/22. pmid: 258
- .
- 2. Миллер К.Д. Канонические вычисления коры головного мозга.Современное мнение в нейробиологии. 2016; 37: 75–84. Epub 2016/02/13. pmid: 26868041; PubMed Central PMCID: PMCPmc4944655.
- 3. Тревес А. Фронтальные защелкивающиеся сети: возможная нейронная основа бесконечной рекурсии. Когнитивная нейропсихология. 2005. 22 (3): 276–91. Epub 2005/01/01. pmid: 21038250.
- 4. Mountcastle VB. Модальность и топографические свойства одиночных нейронов соматической сенсорной коры кошек. Журнал нейрофизиологии. 1957. 20 (4): 408–34. Epub 1957/07/01.pmid: 13439410.
- 5. Lubke J, Feldmeyer D. Поток возбуждающих сигналов и связь в корковом столбце: фокус на коре головного мозга. Структура и функции мозга. 2007. 212 (1): 3–17. Epub 2007/08/25. pmid: 17717695.
- 6. Дуглас Р.Дж., Мартин К.А. Нейронные цепи неокортекса. Ежегодный обзор нейробиологии. 2004. 27: 419–51. Epub 2004/06/26. pmid: 15217339.
- 7. Дуглас Р.Дж., Мартин К.А. Картирование матрицы: пути неокортекса. Нейрон.2007. 56 (2): 226–38. Epub 2007/10/30. pmid: 17964242.
- 8. Haeusler S, Maass W. Статистический анализ свойств обработки информации пластинчатых моделей кортикальных микросхем. Кора головного мозга (Нью-Йорк, Нью-Йорк: 1991). 2007. 17 (1): 149–62. Epub 2006/02/17. pmid: 16481565.
- 9. Зильберберг Г., Гупта А., Маркрам Х. Стереотипия в неокортикальных микросхемах. Тенденции нейробиологии. 2002. 25 (5): 227–30. Epub 2002/04/26. pmid: 11972952.
- 10. Видьясагар TR.Нейронная модель акцента внимания: теменная направляет височное. Исследования мозга Обзоры исследований мозга. 1999. 30 (1): 66–76. Epub 1999/07/17. pmid: 10407126.
- 11. Иса Т., Кобаяши Ю. Переключение между корковыми и подкорковыми сенсомоторными путями. Прогресс в исследованиях мозга. 2004. 143: 299–305. Epub 2003/12/05. pmid: 14653174.
- 12. Джонсон С., Марро Дж., Торрес Дж. Дж. Устойчивая кратковременная память без синаптического обучения. ПлоС один. 2013; 8 (1): e50276. Epub 2013/01/26.pmid: 23349664; PubMed Central PMCID: PMCPmc3551937.
- 13. Шуз А., Брайтенберг В. Белое вещество коры головного мозга человека: количественные аспекты кортико-корковой связи на больших расстояниях. Корковые области: единство и разнообразие. 5. Лондон и Нью-Йорк: Тейлор и Фрэнсис; 2002. с. 377.
- 14. Лопеш да Силва Ф. Х., Хукс А., Смитс Х., Зеттерберг Л. Х. Модель ритмической активности мозга. Альфа-ритм таламуса. Кибернетик. 1974. 15 (1): 27–37. Epub 1974/05/31. pmid: 4853232.
- 15. Фриман WJ. Массовое воздействие на нервную систему. Нью-Йорк: Academic Press; 1975.
- 16. Jansen BH, Rit VG. Электроэнцефалограмма и генерация визуального вызванного потенциала в математической модели связанных столбцов коры. Биологическая кибернетика. 1995. 73 (4): 357–66. Epub 1995/09/01. pmid: 7578475.
- 17. Лили Д.Т., Кадуш П.Дж., Дафилис депутат. Пространственно-непрерывная теория среднего поля электрокорковой активности. Сеть (Бристоль, Англия).2002. 13 (1): 67–113. Epub 2002/03/07. pmid: 11878285.
- 18. Робинсон PA, Ренни CJ, Роу DL. Динамика масштабной мозговой активности при нормальных состояниях возбуждения и эпилептических припадках. Физический обзор E, Статистическая, нелинейная физика и физика мягкой материи. 2002; 65 (4 Пт 1): 041924. Epub 2002/05/15. pmid: 12005890.
- 19. Touboul J, Wendling F, Chauvel P, Faugeras O. Активность нейронных масс, бифуркации и эпилепсия. Нейронные вычисления. 2011. 23 (12): 3232–86. Epub 2011/09/17.pmid: 217.
- 20. Spiegler A, Kiebel SJ, Atay FM, Knosche TR. Бифуркационный анализ моделей нейронной массы: влияние внешних входов и дендритных постоянных времени. NeuroImage. 2010. 52 (3): 1041–58. Epub 2010/01/05. pmid: 20045068.
- 21. Гримберт Ф., Фогерас О. Бифуркационный анализ модели нейронных масс Янсена. Нейронные вычисления. 2006. 18 (12): 3052–68. Epub 21.10.2006. pmid: 17052158.
- 22. Феллеман ди-джей, Ван Эссен, округ Колумбия. Распределенная иерархическая обработка в коре головного мозга приматов.Кора головного мозга (Нью-Йорк, Нью-Йорк: 1991). 1991; 1 (1): 1–47. Epub 1991/01/01. pmid: 1822724.
- 23. Гарнье А., Видаль А., Юно С., Бенали Х. Модель нейронной массы с прямыми и косвенными петлями возбуждающей обратной связи: идентификация бифуркаций и временной динамики. Нейронные вычисления. 2015; 27 (2): 329–64. Epub 2014/12/17. pmid: 25514111.
- 24. Малагаррига D, Вилла AE, Гарсия-Охалво J, Понс AJ. Мезоскопическое разделение возбуждения и торможения в сетевой модели мозга.Вычислительная биология PLoS. 2015; 11 (2): e1004007. Epub 2015/02/12. pmid: 25671573; Идентификатор PubMed Central PMCID: PMCPmc4324935.
- 25. Рудольф М., Поспишил М., Тимофеев И., Дестекс А. Ингибирование определяет динамику мембранного потенциала и контролирует генерацию потенциала действия в коре головного мозга бодрствующей и спящей кошки. Журнал неврологии: официальный журнал Общества неврологии. 2007. 27 (20): 5280–90. Epub 2007/05/18. pmid: 17507551.
- 26.
Дехгани Н., Пейраш А., Теленчук Б., Ле Ван Куен М., Халгрен Э., Кэш С.С. и др.Динамический баланс возбуждения и торможения в неокортексе человека и обезьяны. Научные отчеты. 2016; 6: 23176. Epub 2016/03/17. pmid: 26980663; Идентификатор PubMed Central PMCID: PMCPmc47
. - 27. Цибуркус Дж., Крессман Дж. Р., Шифф С. Дж. Припадки как состояния дисбаланса: возбуждающая и тормозящая проводимость во время припадков. Журнал нейрофизиологии. 2013. 109 (5): 1296–306. Epub 2012/12/12. pmid: 23221405; PubMed Central PMCID: PMCPmc3602838.
- 28. Ваттикути С, Чау СС.Вычислительная модель дисфункции коры головного мозга при расстройствах аутистического спектра. Биологическая психиатрия. 2010. 67 (7): 672–8. Epub 2009/11/03. pmid: 19880095; Идентификатор PubMed Central PMCID: PMCPmc3104404.
- 29. Буржерон Т. Синаптический путь к аутизму. Современное мнение в нейробиологии. 2009. 19 (2): 231–4. Epub 2009/06/24. pmid: 19545994.
- 30. Gogolla N, Leblanc JJ, Quast KB, Sudhof TC, Fagiolini M, Hensch TK. Дефект общей цепи возбудительно-тормозного баланса в мышиных моделях аутизма.Журнал нарушений психического развития. 2009. 1 (2): 172–81. Epub 2010/07/29. pmid: 20664807; PubMed Central PMCID: PMCPmc22.
- 31. Ижар О., Фенно Л. Е., Пригге М., Шнайдер Ф., Дэвидсон Т. Дж., О’Ши Д. Д. и др. Неокортикальный баланс возбуждения / торможения при обработке информации и социальная дисфункция. Природа. 2011. 477 (7363): 171–8. Epub 29.07.2011. pmid: 21796121; PubMed Central PMCID: PMCPmc4155501.
- 32. Сигурдссон Т. Дисфункция нервной цепи при шизофрении: выводы на животных моделях.Неврология. 2016; 321: 42–65. Epub 2015/07/08. pmid: 26151679.
- 33. Роуэн М.С., Неймотин С.А., Литтон У. Электростимуляция для уменьшения прогрессирования болезни Альцгеймера, вызванного синаптическим масштабированием. Границы вычислительной нейробиологии. 2014; 8: 39. Epub 2014/04/26. pmid: 24765074; PubMed Central PMCID: PMCPmc3982056.
- 34. ван Фрисвейк C, Сомполинский Х. Хаос в нейронных сетях со сбалансированной возбуждающей и тормозящей активностью. Наука (Нью-Йорк, Нью-Йорк).1996. 274 (5293): 1724–6. Epub 1996/12/06. pmid: 8939866.
- 35. Шу Y, Hasenstaub A, McCormick DA. Включение и выключение периодической сбалансированной корковой активности. Природа. 2003. 423 (6937): 288–93. Epub 2003/05/16. pmid: 12748642.
- 36. Хайдер Б., Дуке А., Хазенштауб А. Р., Маккормик Д. А.. Активность неокортикальной сети in vivo создается за счет динамического баланса возбуждения и торможения. Журнал неврологии: официальный журнал Общества неврологии.2006. 26 (17): 4535–45. Epub 2006/04/28. pmid: 16641233.
- 37. Бастос А.М., Литвак В., Моран Р., Босман К.А., Фрис П., Фристон К.Дж. DCM-исследование спектральной асимметрии в связях прямой и обратной связи между визуальными областями V1 и V4 у обезьяны. NeuroImage. 2015; 108: 460–75. Epub 2015/01/15. pmid: 25585017; Идентификатор PubMed Central PMCID: PMCPMC4334664.
- 38. Дэвид О., Кибель С.Дж., Харрисон Л.М., Мэттаут Дж., Килнер Дж. М., Фристон К.Дж. Динамическое причинно-следственное моделирование вызванных ответов на ЭЭГ и МЭГ.NeuroImage. 2006. 30 (4): 1255–72. Epub 2006/02/14. pmid: 16473023.
- 39. Jansen BH, Zouridakis G, Brandt ME. Математическая модель вспышек зрительных вызванных потенциалов, основанная на нейрофизиологии. Биологическая кибернетика. 1993. 68 (3): 275–83. Epub 1993/01/01. pmid: 8452897.
- 40. Wendling F, Bartolomei F, Bellanger JJ, Chauvel P. Быстрая эпилептическая активность может быть объяснена моделью нарушенного ГАМКергического торможения дендритов. Европейский журнал нейробиологии.2002. 15 (9): 1499–508. Epub 2002/05/25. pmid: 12028360.
- 41. Гудфеллоу М., Шиндлер К., Байер Г. Самоорганизованные переходные процессы в модели нейронной массы эпилептогенной тканевой динамики. NeuroImage. 2012. 59 (3): 2644–60. Epub 29.09.2011. pmid: 21945465.
- 42. Кунце Т., Хунольд А., Хауэйзен Дж., Джирса В., Шпиглер А. Транскраниальная стимуляция постоянным током изменяет функциональную связность в состоянии покоя: крупномасштабное исследование моделирования сети мозга. NeuroImage. 2016. Epub 2016/02/18.pmid: 26883068.
- 43. Мерле I, Бирот Дж., Сальвадор Р., Молаи-Ардекани Б., Меконнен А., Сориа-Фриш А. и др. От осцилляторной стимуляции транскраниального тока до изменений ЭЭГ кожи головы: биофизическое и физиологическое моделирование. ПлоС один. 2013; 8 (2): e57330. Epub 2013/03/08. pmid: 23468970; Идентификатор PubMed Central PMCID: PMCPmc3585369.
- 44. Меффин Х., Беркитт А.Н., Грейден Д.Б. Аналитическая модель «большого колеблющегося состояния синаптической проводимости», типичного для нейронов неокортекса in vivo.Журнал вычислительной нейробиологии. 2004. 16 (2): 159–75. Epub 2004/02/06. pmid: 14758064.
- 45. Брюнель Н. Динамика слабо связанных сетей возбуждающих и тормозных нейронов. Журнал вычислительной нейробиологии. 2000. 8 (3): 183–208. Epub 2000/05/16. pmid: 10809012.
- 46. Энгельборгс К. Численный бифуркационный анализ дифференциальных уравнений с запаздыванием с использованием DDE-BIFTOOL. Сделки по математическому программному обеспечению. 2002. 28 (1): 1–21.
- 47.Беггс Дж. Гипотеза критичности: как локальные корковые сети могут оптимизировать обработку информации. Философские труды Серия A, Математические, физические и инженерные науки. 2008. 366 (1864): 329–43. Epub 2007/08/04. pmid: 17673410.
- 48. Гессе Дж., Гросс Т. Самоорганизованная критичность как фундаментальное свойство нейронных систем. Границы системной нейробиологии. 2014; 8: 166. Epub 2014/10/09. pmid: 25294989; Идентификатор PubMed Central PMCID: PMCPmc4171833.
- 49.Rolls ET, Deco G. Сети для памяти, восприятия и принятия решений, а также того, как синтаксис языка может быть реализован в мозгу. Исследование мозга. 2015; 1621: 316–34. Epub 2014/09/23. pmid: 25239476.
- 50. Фрейзер Л. О понимании предложений: стратегии синтаксического анализа [докторская диссертация]. докторская диссертация: Университет Коннектикута; 1979.
- 51. Валенсуэла, CF. Взаимодействие алкоголя и нейротрансмиттеров. Здоровье алкоголя и мир исследований.1997. 21 (2): 144–8. Epub 1997/01/01. pmid: 15704351.
- 52. Дуглас Р.Дж., Мартин К.А. Функциональная микросхема зрительной коры головного мозга кошки. Журнал физиологии. 1991; 440: 735–69. Epub 1991/01/01. pmid: 1666655; Идентификатор PubMed Central PMCID: PMCPmc1180177.
- 53. Харрис К.Д., пастырь GM. Неокортикальный контур: темы и вариации. Природа нейробиологии. 2015; 18 (2): 170–81. Epub 2015/01/28. pmid: 25622573; Идентификатор PubMed Central PMCID: PMCPmc4889215.
- 54. Петерс Дж. Ф., Тоцци А., Раманна С.Тесселяция мозговой ткани показывает отсутствие канонических микросхем. Письма о неврологии. 2016; 626: 99–105. Epub 2016/05/26. pmid: 27222926.
- 55. Beul SF, Hilgetag CC. К «канонической» агранулярной кортикальной микросхеме. Границы нейроанатомии. 2014; 8: 165. Epub 2015/02/03. pmid: 25642171; Идентификатор PubMed Central PMCID: PMCPmc4294159.
- 56. Дуглас Р.Дж., Мартин К.А. Рекуррентные нейронные цепи неокортекса. Современная биология: CB. 2007; 17 (13): R496–500. Epub 2007/07/06.pmid: 17610826.
- 57. Carandini M, Heeger DJ. Нормализация как каноническое нейронное вычисление. Обзор Nature Neuroscience. 2011. 13 (1): 51–62. Epub 2011/11/24. pmid: 22108672; Идентификатор PubMed Central PMCID: PMCPMC3273486.
- 58. Haeusler S, Schuch K, Maass W. Распределение мотивов, динамические свойства и вычислительная производительность двух шаблонов кортикальных микросхем на основе данных. Журнал физиологии, Париж. 2009. 103 (1–2): 73–87. Epub 2009/06/09. pmid: 19500669.
- 59. Маасс В., Джоши П., Зонтаг Е.Д. Вычислительные аспекты обратной связи в нейронных цепях. Вычислительная биология PLoS. 2007; 3 (1): e165. Epub 2007/01/24. pmid: 17238280; Идентификатор PubMed Central PMCID: PMCPMC1779299.
- 60. Потянс Т.С., Дисманн М. Кортикальная микросхема, специфичная для клеточного типа: взаимосвязь между структурой и активностью в полномасштабной модели импульсной сети. Кора головного мозга (Нью-Йорк, Нью-Йорк: 1991). 2014. 24 (3): 785–806. Epub 2012/12/04. pmid: 23203991; Идентификатор PubMed Central PMCID: PMCPMC3
8. - 61. Пульвермюллер Ф., Гарагнани М., Веннекерс Т. Мышление схемами: к нейробиологическому объяснению в когнитивной нейробиологии. Биологическая кибернетика. 2014; 108 (5): 573–93. Epub 2014.06.19. pmid: 24939580; Идентификатор PubMed Central PMCID: PMCPMC4228116.
- 62. Веннекерс Т., Гарагнани М., Пульвермюллер Ф. Языковые модели, основанные на сборках геббийских ячеек. Журнал физиологии, Париж. 2006. 100 (1–3): 16–30. Epub 2006/11/04. pmid: 17081735.
- 63. Каин Н., Айер Р., Кох С., Михалас С.Вычислительные свойства упрощенной модели кортикального столбика. Вычислительная биология PLoS. 2016; 12 (9): e1005045. Epub 2016/09/13. pmid: 27617444; Идентификатор PubMed Central PMCID: PMCPMC5019422.
- 64. Фристон К.Дж., Харрисон Л., Пенни В. Динамическое причинно-следственное моделирование. NeuroImage. 2003. 19 (4): 1273–302. Epub 2003/09/02. pmid: 12948688.
- 65. Деку Г., Джирса В.К., Робинсон П.А., Брейкспир М., Фристон К. Динамический мозг: от импульсных нейронов до нейронных масс и корковых полей.Вычислительная биология PLoS. 2008; 4 (8): e1000092. Epub 2008/09/05. pmid: 18769680; Идентификатор PubMed Central PMCID: PMCPmc2519166.
- 66. Бастос А.М., Усрей В.М., Адамс Р.А., Мангун Г.Р., Фрис П., Фристон К.Дж. Канонические микросхемы для предсказательного кодирования. Нейрон. 2012. 76 (4): 695–711. Epub 2012/11/28. pmid: 23177956; Идентификатор PubMed Central PMCID: PMCPMC3777738.
- 67. Босман К.А., Абоитиз Ф. Функциональные ограничения в эволюции мозговых цепей. Границы неврологии. 2015; 9: 303.Epub 2015/09/22. pmid: 26388716; Идентификатор PubMed Central PMCID: PMCPMC4555059.
- 68. Пиноцис Д.А., Шварцкопф Д.С., Литвак В., Рис Г., Барнс Г., Фристон К.Дж. Динамическое каузальное моделирование латеральных взаимодействий в зрительной коре. NeuroImage. 2013; 66: 563–76. Epub 2012/11/07. pmid: 23128079; Идентификатор PubMed Central PMCID: PMCPMC3547173.
- 69. Дюрстевиц Д., Моряки Ю.К., Сейновски Т.Дж. Нейровычислительные модели рабочей памяти. Природа нейробиологии. 2000; 3 Suppl: 1184–91. Epub 2000/12/29.pmid: 11127836.
- 70. Абелес М. Кортиконикс, Нейронные цепи коры головного мозга: Издательство Кембриджского университета; 1991.
- 71. Хебб ДО. Организация поведения: нейропсихологическая теория. Нью-Йорк: Уайли; 1949.
- 72. Ройер С., Мартина М., Паре Д. Бистабильное поведение тормозных нейронов, контролирующих импульсный трафик через миндалину: роль медленно деинактивирующего тока K +. Журнал неврологии: официальный журнал Общества неврологии.2000. 20 (24): 9034–9. Epub 2000/01/11. pmid: 11124979.
- 73. Мардер Э., Гоайярд Дж. М.. Изменчивость, компенсация и гомеостаз нейронных и сетевых функций. Обзор Nature Neuroscience. 2006; 7 (7): 563–74. Epub 2006/06/23. pmid: 167.
- 74. Лю Г. Локальный структурный баланс и функциональное взаимодействие возбуждающих и тормозных синапсов в дендритах гиппокампа. Природа нейробиологии. 2004. 7 (4): 373–9. Epub 2004/03/09. pmid: 15004561.
- 75.Markert H, Knoblauch A, Palm G. Моделирование синтаксической обработки в коре головного мозга. Биосистемы. 2007. 89 (1–3): 300–15. Epub 2007/02/06. pmid: 17276587.
- 76. Friederici AD. К нейронной основе обработки слуховых предложений. Тенденции в когнитивных науках. 2002. 6 (2): 78–84. Epub 2005/05/04. pmid: 15866191.
- 77. Sammler D, Kotz SA, Eckstein K, Ott DV, Friederici AD. Просодия соответствует синтаксису: роль мозолистого тела. Мозг: журнал неврологии.2010. 133 (9): 2643–55. Epub 2010/08/31. pmid: 20802205.
- 78. Марслен-Уилсон В., Тайлер Л.К. Временная структура понимания разговорной речи. Познание. 1980. 8 (1): 1–71. Epub 1980/03/01. pmid: 7363578.
- 79. Йылдыз И.Б., фон Кригштейн К., Кибель С.Дж. От пения птиц до распознавания речи человека: байесовский вывод об иерархии нелинейных динамических систем. Вычислительная биология PLoS. 2013; 9 (9): e1003219. Epub 2013/09/27. pmid: 24068902; Идентификатор PubMed Central PMCID: PMCPmc3772045.
- 80. Пульвермюллер Ф. Взгляд мозга на языковые механизмы: от дискретных нейронных ансамблей до последовательного порядка. Успехи нейробиологии. 2002. 67 (2): 85–111. Epub 2002/07/20. pmid: 12126657.
Модель индивидуализированных канонических микросхем, поддерживающих когнитивные операции
В этом исследовании мы демонстрируем, как локальная архитектура коры головного мозга, представленная канонической микросхемой, реализует основные операции обработки информации стробирования потока сигналов и рабочей памяти.Эти базовые операции формируют основные предпосылки для корковых вычислений более высокого порядка, таких как построение структуры. Мы исследовали, в какой степени локальный топологический выбор ограничивает эти основные операции. Мы продемонстрировали, что только модели с отдельными возбуждающими входными и выходными популяциями (т. Е. Модель трех популяций с косвенной возбуждающей обратной связью) показывают все соответствующие варианты поведения одновременно для одной параметризации. Важно отметить, что баланс локальной сети оказался решающим фактором для доступности основных операций и, следовательно, функциональной индивидуализации микросхемы.Мы продемонстрировали, как сеть, состоящая из нескольких индивидуальных канонических микросхем, может реализовать сложную когнитивную операцию, а именно синтаксический анализ при обработке предложений.
Далее мы обсудим, как наш выбор основных операций и канонической модели микросхемы можно рассматривать как общий знаменатель для различных других утверждений, встречающихся в литературе. Мы будем утверждать, что модели нейронной массы являются подходящим выбором для представления канонической микросхемы, и укажем, как наши исследования выходят за рамки предыдущих исследований моделей нейронной массы.Более того, мы разъясним сильные стороны и ограничения представленного примера в свете нейролингвистики, а также других вычислительных моделей обработки предложений.
Базовые операции в канонических микросхемах
Представление о разнообразных когнитивных функциях, основанных на относительно однородной локальной архитектуре нижележащего нейронного субстрата, поддерживает идею канонических микросхем [6, 52], эффективное взаимодействие которых составляет вычислительную мощность головного мозга. кора [1–3].На данный момент нейробиологические исследования согласуются с такими характеристиками коры головного мозга, как ламинация, биофизические свойства доминирующих типов нейронов, а также целевые и исходные слои передаваемых сигналов [6, 53]. Заметим, однако, что эта концепция не совсем бесспорна [54, 55].
Канонические микросхемы ранее ассоциировались с базовыми операциями или стереотипными функциями [9], такими как управление усилением и восстановление сигнала [56], линейными (например, суммирование, деление и обращение знака) и нелинейными операциями (например.g., «победитель получает все», инвариантность и мультистабильность) [7], усиление и нормализация сигнала [55, 57], а также селективность и вычисление коэффициента усиления [2]. Крупномасштабные импульсные нейронные сети, явно имитирующие ламинарную архитектуру кортикального столба, широко используются для изучения связи между стереотипной структурой и базовыми операциями в вычислительных моделях канонических микросхем [8, 58–60]. Наличие таких операций также исследовалось в моделях среднего поля [61–63], которые составляют основу большинства попыток моделирования нейрокогнитивных экспериментов (например,г., DCM; [37, 64]). В настоящем исследовании мы продвигаем понимание канонических операций, предоставляя доказательства фундаментальных базовых операций даже в очень простой модели обратной связи канонической микросхемы. Из-за своего фундаментального характера идентифицированные базовые операции (стробирование потока сигналов и рабочая память) подразумевают другие ранее описанные канонические операции (см. Выше), которые являются более сложными и были обнаружены в очень специфических архитектурах кортикального столбца. Установлено, что модели среднего поля могут надежно описывать коллективное поведение большого числа нейронов [65].Важно отметить, что пространственная абстрактность выбранного нами типа модели, гибко отражающей взаимодействие нескольких нейронов, нейронных популяций или целых областей коры, позволяет интерпретировать эту базовую функциональность, которая актуальна на различных уровнях нейронной организации, потенциально превышая уровень коркового столба.
Тем не менее, мезоскопический пространственный масштаб нейронных популяций, в котором, возможно, устанавливается однородность канонических микросхем, хорошо отражается концепцией моделей нейронной массы [20, 65].Здесь мы исследуем его динамику в ответ на переходный вход в свете изменений основного пространства состояний, что наиболее важно для обработки информации, управляемой стимулами.
Наше внимание к отдельной микросхеме дополняет последние разработки в области сетевых технологий, которые также обращаются к идее моделирования канонических микросхем с помощью моделей нейронных масс. Было изучено, как очевидное различие межсхемных соединений на прямые и обратные связи, с ламинарно-специфическим происхождением и целями [22, 38, 66], мотивирует расположение канонических микросхем в иерархических моделях [37, 67, 68] .Что еще не изучено в этих моделях, так это основные операции на уровне микросхемы и их чувствительность к топологическим особенностям и уровням возбуждения и торможения. В этом исследовании мы исследовали поведение реакции, вызванной стимуляцией, в трех репрезентативных локальных топологиях: (i) трехпопуляционная модель (), (ii) двухпопуляционная модель () и (iii) двухпопуляционная модель с повторяющимися тормозная обратная связь (). Среди рассмотренных архитектур обратной связи и соответствующих диапазонов параметров мы обнаружили, что только модель с тремя популяциями (с входными данными для возбуждающих интернейронов, таким образом, с отдельными входными и выходными популяциями) демонстрирует сосуществование всех трех вариантов поведения ответа для фиксированного значения баланса сети. .То есть только трехпопуляционная архитектура способна выборочно блокировать, передавать или запоминать стимул в зависимости от его свойств. Кроме того, поведение переноса моделей с двумя популяциями зависит только от силы применяемых стимулов, а не от их продолжительности, в отличие от модели с тремя популяциями, где оно зависит от обоих. В этом смысле наши результаты демонстрируют, что рассмотрение косвенной возбуждающей петли обратной связи очень важным образом увеличивает разнообразие и биологический реализм динамики модели.Поскольку более сложные архитектуры моделей обычно демонстрируют более богатый динамический репертуар, мы ожидаем, что такое поведение также будет иметь место в более детальных моделях канонических микросхем. В частности, наша модель не учитывает прямое взаимодействие между возбуждающими и тормозными интернейронами, а также саморегуляторную связь между возбуждающими интернейронами. Обе эти связи, безусловно, существуют [58]. Эти особенности являются одними из важных расширений модели, которые необходимо изучить в будущих исследованиях.
В нашей модели канонической микросхемы мы описали поведение локальной памяти на основе бистабильности. Повторяющаяся и самоподдерживающаяся активность модулируется балансом сети и инициируется и прекращается афферентными входами, поведение, которое было показано in vitro [35]. В отличие от долговременной памяти, которая опирается на синаптическую пластичность, краткосрочная (или рабочая) память, как полагают, полагается на механизмы, которые не изменяют основную структуру связности. В качестве альтернативы было предложено, что рабочая память может быть основана на задержках и постоянных времени [12, 69], таких как цепочки синфайров , [70], повторяющиеся возбуждающие сети [71] или свойства клеток [72].В этих механизмах период хранения (время забывания) зависит от относительно фиксированных структурных аспектов сети, тогда как при бистабильности элемент может храниться в памяти в принципе в течение любого периода, пока он не будет активно выключен (например, с помощью импульса ингибирующая популяция, см. S2 фиг.).
Нейронные функции должны быть устойчивы к шуму, но в то же время чувствительны к афферентному входу. Сигналы можно отличить от шума по более высокой амплитуде и плавности во времени.В нашей модели по амплитуде расстояние между рабочей точкой и бифуркацией складки должно быть больше шума и меньше сигнала. Мы продемонстрировали, что это расстояние эффективно контролируется балансом локальной сети, который, следовательно, является ключевым параметром для управления компромиссом между устойчивостью к шуму и чувствительностью к стимулам (например, S4 Fig). Что касается временной гладкости, только трехпопуляционная модель предлагает механизм отбора. Мы установили, что модель сохраняет стимул в памяти только в том случае, если он длится достаточно долго, то есть демонстрирует большую степень временной плавности.
В дополнение к концепции функциональной диверсификации по топологии [3], мы показали, что изменение баланса локальной сети также является биологически правдоподобным средством индивидуализации локальной функциональности. Эта локальная настройка определяет, будет ли нейронная область выборочно пересылать импульс (поведение передачи), переключаться на более высокую постоянную активацию (поведение памяти) или вообще не реагировать на импульс (невосприимчивое поведение). Отметим, что в этом случае мы рассматривали баланс локальной сети как сосредоточенный параметр, который фактически инкапсулирует большое количество физиологических и анатомических свойств и механизмов, таких как кинетика нейротрансмиттеров и плотности нейрорецепторов, дендритное ветвление, синаптическая и внеклеточная ионная динамика и подключения к локальной и нелокальной сети, и это лишь некоторые из них [73, 74].Из-за этой двусмысленности и, как обсуждалось ранее, концепцию баланса сети очень трудно дать количественной оценке. В этом исследовании мы использовали простое определение, чтобы продемонстрировать в качестве доказательства концепции, что баланс локальной сети действительно обеспечивает фундаментальное средство индивидуализации функциональности локальной микросхемы.
Применение канонической микросхемы для обработки предложений
Понятие обширной сети подобных микросхем, поддерживающих корковые функции, было выдвинуто в специализированных нейрокогнитивных теориях.Например, было выдвинуто несколько предложений, чтобы обеспечить механистическое понимание обработки языка [1, 49, 61, 62, 75]. Это также было связано с организационными принципами, такими как кодирование места, где нейронная область представляет собой абстрактный элемент обработки [49]. В этом исследовании мы использовали канонические микросхемы для представления составляющих воспринимаемых предложений, отражая принцип кодирования мест. Канонические микросхемы, представляющие одно слово, были сгруппированы в модули в соответствии с их синтаксической ролью (например,g., субъект, объект), а не их словесную категорию (например, существительное, местоимение). Мы описываем вычислительный механизм управляемой вводом функциональной привязки дискретных элементов. Это позволяет генерировать бесконечные последовательности из ограниченного числа дискретных элементов, концепция, называемая бесконечной рекурсией [3]. Предлагаемый механизм построения генеративной структуры можно было бы назвать динамическим набором , чтобы подчеркнуть свободу структуры в отношении ее количества и порядка элементов.
Предлагаемый вычислительный механизм применяется для синтаксического анализа, то есть группирования непрерывного потока слов в иерархическую структуру составных частей предложения. Мы ограничиваем механизм построения структуры, фиксируя лежащую в основе проводку взаимодействующих нейронных областей. Характеристики сети, то есть ее топология и веса соединений, отражают правила синтаксиса, которые, вероятно, будут установлены во время изучения языка. Базовые операции, обеспечиваемые канонической микросхемой, имеют решающее значение для этого механизма построения структуры.Очевидно, что для реализации временной интеграции последовательностей в реальном времени необходим эффективный механизм краткосрочной памяти. Механизм рабочей памяти нашей канонической микросхемы обеспечивает быстрое кодирование и гибкое время хранения. Последнее особенно актуально для учета разной скорости речи и разной длины предложений. Стробирование потока сигналов гарантирует, что слова активируются только в том случае, если они были предварительно активированы (ожидаемы) структурой предложения и распознаны из входного потока.Поскольку было показано, что эти базовые операции в значительной степени зависят от баланса сети, изменение этого параметра приводит к сбою работы глобальной сети. В текущей реализации языковой сети сосуществование памяти и поведения передачи, которое является отличительной чертой модели трех популяций, не является необходимым ингредиентом. Таким образом, также могла быть использована модель двух популяций. Однако для дальнейшей разработки модели могут стать актуальными специфические свойства модели трех популяций.Например, из-за неспецифической информации афферентного слова отдельные слова в принципе могут встречаться несколько раз (например, слово пить как субъект, глагол, объект). Чтобы предотвратить одновременную устойчивую активацию нескольких канонических микросхем (которая приведет к насыщению узлов и сделает их недоступными для последующей активации), локальная схема должна иметь поведение передачи, обеспечиваемое моделью трех популяций.
Предлагается, чтобы в процессе понимания предложения начальный синтаксический анализ, рассматриваемый в данном исследовании, сопровождался тематическим распределением ролей и коллективной оценкой синтаксической и семантической информации [76].Помимо прагматической информации, предлагается также просодическая информация для руководства пониманием предложения [77]. В нашей модели мы объединяем все типы такой дополнительной информации в более общее понятие контекстной информации.
Предлагаемая сетевая модель отражает характеристики serial , модели с первым синтаксисом путем построения простейшей синтаксической структуры на основе информации категории слова, независимой от лексико-семантической информации [76]. Модель также отражает характеристики моделей удовлетворения ограничений [78] за счет включения неструктурных факторов в топологию сети.К таким факторам относятся частота конкретной структуры или ее семантическая правдоподобность. В соответствии с концепцией моделей удовлетворения ограничений предложенная сетевая модель способна учитывать синтаксические неоднозначности (см.). Мы также предполагаем, что восприятие и идентификация словесной информации информирует, но концептуально отличается от вычислений построения структуры. Входящая слуховая информация распознается как слово [79], которое затем вводится в нейронные области, которые задействуются во время синтаксического анализа.
В дополнение к аналогичной модели [49] наша модель рассматривает модификаторы глаголов и объектов. Однако в предложенном здесь механизме построение структуры все еще сильно опирается на временные порядки слов, то есть субъект-глагол-объект. Помимо информации о категории слов, успешный синтаксический анализ также должен учитывать морфосинтаксические особенности, такие как пол, префиксы или падежи. Каким образом эти функции встроены — вопрос открытый и требует дальнейшего изучения.
Одной из целей нашего исследования было проложить путь к механистическому пониманию обработки предложений в соответствии с нейропсихологическими теориями.Это противоположно области компьютерной лингвистики, которая направлена на оптимизацию разговорного взаимодействия человека с машиной, жертвуя при этом биологической достоверностью. Кроме того, мы хотели бы подчеркнуть характер развития нашей модели и осознаем несколько недостатков, некоторые из которых мы рассматриваем здесь:
На уровне слова
Трудно справиться с множественными упоминаниями одного и того же слова в предложении. с. Как только слово в отдельном модуле активируется, его необходимо деактивировать, прежде чем его снова можно будет активировать.Хотя механизм деактивации неявно включен в нашу модель, через короткий импульс к IIN, деактивация в режиме онлайн одного слова прервала бы репрезентативную трассировку активации. Кроме того, суммирующий эффект нескольких предварительных активаций в конечном итоге приведет к активации области, даже если слово отсутствует на входе. Расширение модели могло бы масштабировать уровень предварительной активации относительно порога активации, так что слово «на кончике языка», но еще не активировано.
На уровне предложения
Опять же, представления необходимо деактивировать, прежде чем можно будет представить следующее предложение. Чтобы следить за беседой, необходимо активировать последовательность слов и связанные с ними словесные сети [80]. Эти сети медленно распадаются, так что информация, охватывающая несколько предложений, может быть связана вместе посредством ассоциативных процессов. Наконец, ранее было показано, что сети со слишком большим количеством активированных узлов имеют тенденцию становиться нестабильными и тем самым уничтожать информацию, хранящуюся в состоянии сети [42], которая может иметь отношение к длинным или сложным предложениям.
Появление когнитивного прайминга и построения структуры на основе иерархического взаимодействия канонических моделей микросхем
Далее мы сначала продемонстрируем, что модель mCMC различает афферентную прямую и обратную связь, а также то, как информация обратной связи обуславливает доступность базовых операций обработки. Затем мы покажем, как идентифицированный сигнал стимулирующей обратной связи приводит к более высокой обработке информации, а именно к первичной обработке и построению структуры, в прототипных метасхемах двух взаимодействующих mCMC.Наконец, мы применяем наши выводы к языковой сети для синтаксического анализа, которая объединяет синтаксические предсказания и сенсорную информацию о словах для создания прототипа синтаксиса.
Входные каналы различают информационный поток
Предлагаемая модель mCMC включает две отдельные группы для получения информации с прямой связью и обратной связи соответственно. Мы стимулировали обе популяции независимо друг от друга прямоугольными импульсами определенной величины и продолжительности и нанесли на карту соответствующее поведение реакции (отсутствие реакции, передача и память; см. «Теория и анализ»).
Для стимуляции Py мы наблюдали большие диапазоны невосприимчивого поведения и только редкие следы поведения памяти, встроенные в переходное поведение (рис. 3b). Напротив, те же самые стимулы, примененные к EIN, вызывали невосприимчивое поведение для слабых стимулов, переходное поведение для сильных, но кратких стимулов и поведение памяти для сильных и длительных стимулов (рис. 2b) (Kunze et al., 2017). Это подтверждает движущий характер входов с прогнозированием. Обратите внимание, что полосатая реакция памяти отражает зависимость фазы собственных колебаний системы от времени выключения стимула, что дополнительно объясняется в Kunze et al.(2017).
Рис. 3Модулирующее влияние одновременной стимуляции ЭИН и Py. a Для постоянных уровней p Py и p EIN , двухпараметрическая бифуркационная диаграмма отображала ход бифуркаций складки и Хопфа (цветные линии). Цветными поверхностями отмечены входные конфигурации, для которых присутствовали моностабильные (серые и желтые оттенки) или бистабильные состояния (зеленые оттенки). b — e В отличие от рис.2, мы применяли возрастающие уровни постоянного входного сигнала к EIN одновременно с временной стимуляцией Py. b Без модулирующего входа характерный отпечаток пальца (справа) показывает только редкие следы поведения памяти (оранжевый). c — e Для более высоких уровней модуляции ввода p EIN характерные отпечатки пальцев (справа) показывают увеличенные диапазоны стимуляции, которые вызывают поведение памяти. Это происходит из-за исчезновения сепаратрисы, которая ограничивает поведение памяти (левые проекции пространства состояний).В отпечатках пальцев пороговое значение интенсивности, которое отделяло неотзывчивое (зеленый) и переходное (серый) поведение, уменьшилось, потому что расстояние между рабочей точкой ( p Py = 0) и бифуркацией нижней складки уменьшилось (цветной рисунок онлайн)
Эти отклики поведение было основано на динамике в пространстве состояний. Для обеих целей стимуляции (EIN и Py) анализ бифуркации и устойчивости дал S-образную кривую с фиксированной точкой, поворотные точки которой отражают бифуркации складок, одна типа седло-седло и одна типа седло-узел (левые панели на рис.2б, 3б). Два стабильных участка кривой с фиксированной точкой (один с более высокой и один с более низкой активностью, см. Сплошные линии на левой панели) обозначают бистабильность, которая обуславливает базовую работу рабочей памяти. Расстояние между рабочей точкой ( p EIN = p Py = 0) и бифуркацией седло-узел (нижняя складка) отражает порог интенсивности (зелено-серая граница на правой панели на рис. 2b, 3b), который отделяет поведение передачи и памяти от поведения без ответа.Для обеих целей стимуляции субкритическая бифуркация Хопфа приводит к возникновению сепаратрисы, которая ограничивает область притяжения верхнего (запоминающего) состояния. Только если состояние системы находится в пределах этого бассейна в момент выключения входного сигнала, стимул будет запомнен. Таким образом, сигналы стимуляции будут выбираться в соответствии с их временной согласованностью и продолжительностью. Вместе с порогом интенсивности этот механизм фильтра обуславливает операцию стробирования потока сигнала mCMC (см. Kunze et al.2017) для получения дополнительной информации о механизмах, лежащих в основе основных операций канонической модели микросхемы). Более того, два стабильных предельных цикла вводят устойчивые колебания для стимуляции Py (левая панель на фиг. 3b), в отличие от стимуляции EIN (левая панель на фиг. 2b). Эти колебания большой амплитуды препятствуют установлению системы внутри сепаратрисы по окончании стимуляции ( p Py = 0). Это объясняет немногочисленные редкие следы поведения реакции памяти в характерном отпечатке пальца (правая панель на рис.3б). Для получения дополнительной информации о бифуркационной структуре системы см. Kunze et al. 2017 (стимуляция EIN) и Spiegler et al. 2010 (стимуляция Py).
Таким образом, поведение отклика указывает на различимые операционные роли для отдельных входных каналов: в то время как оба канала пропускают информацию через пороговое значение интенсивности, только входной канал EIN, который, как предполагается, принимает прямой ввод, позволяет хранить информацию. Эта дифференцированная обработка документирует способность модели по-разному реагировать на афферентные информационные потоки, как это требуется в иерархических установках.
Сопутствующая стимуляция модулирует поведение ответа
Иерархически взаимодействующие мСМЦ могут одновременно получать информацию прямого и обратного характера, эффективно интегрируя сенсорную информацию низкого уровня и концептуальную информацию высокого уровня. Соответственно, мы исследовали временную стимуляцию EIN с прямой связью при одновременном применении постоянного входного сигнала обратной связи, отражающего длительное время удержания из-за более медленных процессов верхнего уровня (Bastos et al.2015), к Py модели mCMC (см. Рис.2а).
Мы задокументировали последующее изменение поведения реакции, вызванной стимуляцией, с помощью бифуркационных диаграмм и характерных отпечатков пальцев (рис. 2). При увеличении уровней постоянного ввода обратной связи на Py и переходного прямого ввода на EIN диапазон запоминаемых стимулов (оранжевая область на рис. 2b – f) увеличивается. Между тем, диапазон стимулов, вызывающих невосприимчивое поведение (зеленая область на рис. 2b – f), уменьшается из-за более низкого порога интенсивности стимуляции (то есть верхнего края зеленой области).Следовательно, вход в Py заставляет воспринимать или даже запоминать ранее не реагирующие стимулы (см. Маркеры на рис. 2) — поведение, подтверждающее модулирующий характер информации обратной связи. Эти изменения в поведении переноса вызваны измененным пространством состояний: согласно бифуркационным диаграммам (левые графики на рис. 2b – f), возрастающий сопутствующий вход Py сдвигает нижнюю бифуркацию кратности к меньшим значениям (с p EIN = 78 с −1 до p EIN = 48 с −1 ) и расширяет нестабильный цикл Хопфа (т.е.е., сепаратриса). Это более короткое расстояние между рабочей точкой ( p EIN = 0) и бифуркацией нижней складки, определяющей порог интенсивности стимуляции, и большее пространство, окруженное сепаратрисой, эффективно сокращая необходимую продолжительность стимуляции для оседания в бассейне привлечение верхней фиксированной точки, способствовать поведению реакции памяти.
Для сравнения мы также рассмотрели менее правдоподобный противоположный случай и применили увеличивающиеся уровни постоянного входного сигнала к EIN одновременно с переходной стимуляцией Py.Мы обнаружили модулирующий эффект, который способствует формированию полосатых реакций памяти, которые очень чувствительны к вариациям продолжительности стимуляции (см. Рис. 3b – e). Следовательно, существует асимметрия во взаимном модулирующем влиянии входных каналов. Базовая структура бифуркации в пространстве состояний дополнительно проясняет эту асимметрию и выявляет конфигурации постоянного входа, для которых фактически присутствует бистабильное поведение, необходимое условие для поведения памяти (см.рис.3а).
Таким образом, одновременная стимуляция модели mCMC прямой и обратной информацией вызывает асимметричную взаимную модуляцию поведения реакции, которая обогащает динамический репертуар. Вход обратной связи в Py эффективно модулирует чувствительность системы к управляющему входу с прямой связью, приложенному к EIN.
Важно отметить, что этот модулирующий эффект вызывает различное поведение реакции на идентичные стимулы и обозначает вход обратной связи как сигнал содействия .Этот стимулирующий сигнал обратной связи можно использовать двумя способами для изменения поведения реакции, вызванной стимуляцией: (i) невоспринимаемый стимул (сравните треугольники на рис. 2b, f) становится воспринимаемым и (ii) либо невоспринимаемым (кружок ) или незапоминаемый стимул (крестик) становится запоминающимся (рис. 2б, е).
Прототипные метасхемы
До сих пор мы показали, как минимальная каноническая модель схемы по-разному обрабатывает информационные потоки с прямой связью и обратной связью, и что ввод обратной связи может эффективно регулировать доступ к базовым операциям для прямого ввода.Далее мы вывели прототипные метасхемы двух взаимодействующих mCMC, которые эффективно используют эти особенности обработки для поддержки прайминга и построения структуры.
В начальной мета-схеме (рис. 4a) mCMC A 1 более высокого уровня передает сигналы вспомогательной обратной связи на mCMC A 2 более низкого уровня, который также получает стимуляцию с прямой связью p ff-Stim . Сигнал обратной связи слабый, если A 1 находится в состоянии низкой активности, и высокий, если A 1 находится в состоянии высокой активности.Эта начальная метасхема представляет собой минимальную каноническую архитектуру, которая устанавливает общий механизм для зависимой от состояния обработки, где активность одного mCMC (здесь A 1 ) управляет индуцированным стимуляцией ответом другого mCMC (здесь A ). 2 ) с очевидными последствиями для восприятия и памяти. Примечательно, что эта зависящая от состояния операция обработки формирует потенциальный строительный блок для прогнозирующего кодирования путем интеграции информации концептуальной модели (ввод обратной связи) с новой сенсорной информацией (ввод с прямой связью). {*} \).{*}, \) может также изменять порог восприятия еще одной цепи. c Облегчение модифицирует запоминание (построение структуры), где \ (\ tilde {A} _ {2} \) считается высшей микросхемой, но все же получает вспомогательный сигнал обратной связи, p fac, в , от \ (\ tilde {A} _ {1} \). Сигнал обратной связи обуславливает запоминание стимуляции с прямой связью p ff-Stim , поступающей от нижнего mCMC \ (\ tilde {A} _ {3} \). В случае такой активации \ (\ tilde {A} _ {2} \) вспомогательный сигнал обратной связи p fac, out передается в подключенные схемы, эффективно каскадируя эту локальную операцию и поддерживая инкрементное наращивание. набор устойчивых паттернов активности
Из этой начальной мета-цепи мы получили прототипную метасхему для прайминга, введя прямое соединение от нижнего к верхнему mCMC (рис.4б). Внутренний механизм, адаптивно смещающий порог восприятия, подробно изучается для его граничных условий в Разд. 3.4.
Чтобы получить прототипную метасхему для построения структуры (рис. 4c), мы концептуально рассматриваем \ (\ tilde {A} _ {2} \) теперь как микросхему более высокого уровня, которая все еще получает сигнал вспомогательной обратной связи p fac, в из \ (\ tilde {A} _ {1} \). Предполагается, что сигнал стимуляции с прямой связью p ff-Stim поступает от другого mCMC нижнего уровня \ (\ tilde {A} _ {3} \), который, в свою очередь, получает стимуляцию для своего EIN.Сигнал стимулирующей обратной связи p fac, в будет эффективно регулировать доступность рабочей памяти в \ (\ tilde {A} _ {2} \), механизме, изученном в разд. 3.5. Предоставляя вспомогательный сигнал p fac, out для дополнительных подключенных схем, эта метасхема построения структуры поддерживает создание пространственно-временных шаблонов активации и используется в качестве модуля для сети синтаксического анализа, рассмотренной в разд. 3.6. Обратите внимание, что в реальных корковых цепях, вероятно, будут также обратные связи от \ (\ tilde {A} _ {2} \) к \ (\ tilde {A} _ {3} \), которые влияют на перенос и запоминание поведения последние и влияют на p ff-Stim .{*} \) не требует поддержки той же схемы, которая получила праймер, т. е. самовсасывания, но может также облегчить обработку в другой микросхеме, принимающей цель, представляя восприятие другой модальности (как показано на рис. 4b ).
Условия существования этого механизма заливки мы охарактеризовали путем более внимательного изучения нескольких основных параметров.
Прирост межсхемной связи c f и c b Сила прямого и обратного соединений масштабирует мощность сигнала на целевых микросхемах.{*} \), вызывая стимулирующий сигнал обратной связи. Мы исследуем взаимосвязь между целевым и первичным стимулами с точки зрения различной интенсивности, сохраняя при этом их длительность одинаковой.
Индивидуальная адаптация микросхем В более раннем исследовании мы показали, что отдельные уровни баланса локальной сети могут влиять на динамику ответа мКМЦ (Kunze et al., 2017). В частности, небольшое увеличение ингибирования по сравнению с параметризацией по умолчанию способствует поведению передачи, тогда как небольшое уменьшение ингибирования способствует поведению памяти.{*} \), и (б) целевой стимул вызвал кратковременную высокую активацию только после прайминга. c Применяли целевые стимулы разной продолжительности ( t dur ) и интенсивности. Сумма всех эффективных потоков стимуляции масштабировалась по общему количеству рассматриваемых потоков стимуляции. d Этот процент с цветовой кодировкой был сопоставлен с диапазоном увеличения связности c f и c b , чтобы охарактеризовать их ограничивающее влияние на механизм заливки
Таблица 2 Значения параметров для оценки перцепционный запорный механизм Рис.7Влияние увеличения количества подключений, интенсивности прайминга и баланса сети на механизм прайминга. Возможности подключения ограничивают потоки эффективной стимуляции. Каждый график варьируется в зависимости от прироста связности c f и c b и суммируется по стимуляциям разной продолжительности и интенсивности (см. Рис. 6). Цвета кодируют процент эффективных потоков стимуляции, вызвавших прайминг. Прайминг и целевой стимул были одинаково продолжительными, но интенсивность праймера была на 20% (левый столбец) или на 80% (правый столбец) больше.{*} \) ( b ) увеличивает максимальную скорость эффективных потоков стимуляции и диапазон подходящих улучшений связности. Обратите внимание на различное цветовое масштабирование на отдельных подзаголовках и на то, что проценты являются специфическими для выбранных диапазонов параметров стимуляции (см. Таблицу 2).
Прирост связности c f и c b ограничивают механизм прайминга (см. {*} \) в схему более высокого уровня во время заливки, и для c b <45, что означает максимальную силу обратной связи, чтобы ограничить содействие в цепи нижнего уровня и сохранить ее чувствительность к дальнейшей стимуляции (т.е., предотвращать самоактивацию). Обратите внимание, однако, что точные значения зависят от выбранной параметризации модели. Эти ограничения связности наблюдались для различной интенсивности праймингового стимула (20% и 80%, соответственно) и для всех рассмотренных конфигураций тормозных синаптических усилений H i (рис. 7). Процент эффективных потоков стимуляции увеличивался с относительной силой прайминга и целевого стимула для всех рассмотренных конфигураций тормозных синаптических усилений H i .{*} \) максимально увеличили диапазон подходящих значений связности и скорость эффективных потоков стимуляции (рис. 7b). Далее мы идентифицировали те целевые стимулы, которые вызывают сдвиг порога восприятия, характерный для механизма прайминга (см. {*} \) (голубой цвет).Настройка тормозящего синаптического усиления увеличила диапазон подходящих параметров стимуляции. Важно отметить, что описанный механизм загрузки недостижим в альтернативных топологиях локальной сети (см. Рис. 9).
Рис. 8Характеристика стимулов, вызвавших смещение порога восприятия. Каждый подзаголовок варьируется в зависимости от продолжительности и интенсивности стимула и показывает количество эффективных стимулов (суммированных, прежде всего, c f — c b комбинаций) в оттенках серого.{*} \) не только способствовали количеству воспринимаемых целевых стимулов, но также их инвариантности к вариациям интенсивности или продолжительности, отмеченным более широким диапазоном эффективных стимулов. Обратите внимание на разные оттенки серого для каждого участка.
Рис. 9Важность топологии прототипа метасхемы. a Для механизма заправки мы рассмотрели конкретную топологию в прототипной метасхеме, где более высокая микросхема подключена к более низкой микросхеме через обратное соединение, т.е.е., нацеливаясь на Py, а нижняя микросхема подключается к вышестоящей микросхеме через прямое соединение, то есть нацеленная на EIN. Такая компоновка поддерживает функциональную специализацию: прямое соединение позволяет хранить события и поддерживает память в вышестоящей схеме. Обратное соединение позволило нижней микросхеме выполнять динамическое стробирование потока сигналов. Напротив, альтернативные топологии, такие как рассмотрение чистой прямой связи ( b ), чистой обратной связи ( c ) или перестановка прямой связи и обратной связи ( d ), не смогли поддержать механизм заливки, но могут быть актуальным для других совместных нейронных операций
Таким образом, мы предлагаем механизм прайминга в прототипной мета-цепи двух мСМЦ, который основан на эффекте упрощения сверху вниз.Механизм основан на конструктивном взаимодействии задействованных мМКЦ и предсказывает их концептуальную специализацию либо на сигнал, стробирующий , тем самым избегая нечувствительности к будущим стимулам, либо на запоминание , чтобы направлять дальнейшую обработку информации посредством фасилитации. Эта функциональная специализация может быть искажена за счет местного соотношения возбуждения и торможения в задействованных микросхемах. Топология прямых и обратных связей внутри мета-цепи и значимость стимулов ограничивают механизм прайминга.
Мета-схема построения структуры
В предыдущем разделе мы показали, как нисходящая фасилитация через обратную связь позволяет динамическое восприятие подсознательных стимулов. Далее мы покажем, как сигнал фасилитации обусловливает запоминание событий стимуляции в мета-цепи построения структуры (рис. 4c).
В метасхеме для построения структуры мы применили переходные прямоугольные стимулы с прямой связью к \ (\ tilde {A} _ {3} \) и отобразили последующие функциональные состояния \ (\ tilde {A} _ {2} \ ) и \ (\ tilde {A} _ {3} \) для увеличения уровней стимулирующей внешней обратной связи p fac, in (см. рис.10). Для подавляющего синаптического усиления по умолчанию слабые и короткие стимулы не могут активировать \ (\ tilde {A} _ {2} \) ( без запоминания , серая область), а сильные и длинные стимулы активируют оба \ (\ tilde { A} _ {2} \) и \ (\ tilde {A} _ {3} \) (, общее запоминание , черная область). Немногие стимулы могут избирательно активировать \ (\ tilde {A} _ {2} \), но избегают устойчивой высокой активности \ (\ tilde {A} _ {3} \) и, таким образом, сохраняют его реакцию на дальнейший ввод ( запоминание и отзывчивость , зеленая зона).Более высокие уровни сигнала стимулирующей обратной связи p fac, in способствуют избирательной активации \ (\ tilde {A} _ {2} \) (красные кресты на фиг. 10a). Небольшое усиление ингибирования в \ (\ tilde {A} _ {3} \) дополнительно способствует этой избирательной активации (см. Рис. 10b), эффективно способствуя поведению реакции передачи (Kunze et al. 2017). Таким образом, нисходящий сигнал содействия p fac, в обуславливает установление устойчивой активности в одной части (т.е.е., \ (\ tilde {A} _ {2} \)) прототипной метасхемы для построения структуры, сохраняя при этом ее входную чувствительность (т.е. \ (\ tilde {A} _ {3} \)). Далее мы проиллюстрируем, как эта локальная операция поддерживает последовательное и выборочное установление пространственно-временных структур в простой реализации инкрементного (то есть пословного) синтаксического синтаксического анализа во время восприятия предложения.
Рис. 10Облегчающий сигнал позволяет запоминать события стимуляции. MCMC \ (\ tilde {A} _ {3} \) получал стимулы разной длины и интенсивности и, в свою очередь, стимулировал mCMC \ (\ tilde {A} _ {2} \) через прямое соединение. a Для подавляющего синаптического усиления по умолчанию несколько стимулов к \ (\ tilde {A} _ {3} \) позволяют избирательную активацию \ (\ tilde {A} _ {2} \) при сохранении \ (\ tilde { A} _ {3} \) отзывчивый (т.е. запоминание и отзывчивость, зеленая зона). Слабые и краткие стимулы не активируют \ (\ tilde {A} _ {2} \) ( без запоминания , серая область), тогда как сильные и длинные стимулы активируют оба \ (\ tilde {A} _ {2} \) и \ (\ tilde {A} _ {3} \) ( общее запоминание , черная область). Более высокие уровни сигнала стимулирующей обратной связи p fac, in способствуют избирательной активации \ (\ tilde {A} _ {2} \). b Небольшое увеличение тормозящего синаптического усиления H i , благоприятствующее поведению реакции передачи в \ (\ tilde {A} _ {3} \), способствует избирательной активации \ (\ tilde {A} _ { 2} \). Красные крестики обозначают примерную стимуляцию определенной интенсивности и продолжительности, применяемую к \ (\ tilde {A} _ {3} \) (цветной рисунок онлайн)
Синтаксическая языковая сеть
Понимание устного или письменного предложения включает множество этапов обработки , включая акустическое восприятие, отображение синтаксической и семантической информации и включение дополнительной информации (т.е., контекст и индивидуальный опыт) (Friederici, 2002). Этот процесс восприятия предложения проявляется как последовательная активация нейронных популяций в задействованных неокортикальных областях (Kunze et al., 2017; Rolls and Deco 2015; Pulvermüller et al. 2014) . В следующем примере мы еще раз возвращаемся к языковой модели (Kunze et al., 2017), которая фокусируется на представлении синтаксической информации в распределенной сети mCMC. В частности, он учитывает декодирование и временное хранение синтаксической информации, здесь именуемой , синтаксический анализ , которая необходима для дальнейшей обработки (Friederici 2002).Считается, что слова представлены сетью слов, состоящей из множества узлов для всех аспектов слова, включая словоформы, различные семантические ассоциации и одну или несколько синтаксических ролей. Здесь мы принимаем упрощенную модель, содержащую для каждого слова только узлы для возможных синтаксических ролей (например, субъект, глагол и объект), которые следуют принципу кодирования места (Rolls and Deco 2015), и один узел для остальных слова Интернет. Каждый узел представлен mCMC.
Последовательная информация афферентных слов, исходящая из слуховых областей коры головного мозга, постепенно вызывает устойчивую более высокую активацию в соответствующих микросхемах, представляющих слово.Этот общий механизм построения структуры, называемый динамическим набором , подчеркивает потенциальную свободу синтаксического анализа в отношении количества и порядка его элементов (Kunze et al., 2017). Анализ этой исходной модели выявил следующие проблемы:
Множественные экземпляры отдельных слов Из-за единого объектного модуля, включающего все известные существительные словаря отдельного человека, исходная модель не могла представить повторное воплощение одного и того же слова, как в предложении «Я рисую стену» на стене ».Распознанное слово активировало соответствующую микросхему, которая стала нечувствительной к дальнейшей стимуляции и, следовательно, была недоступна для дальнейшего набора.
Простая синтаксическая структура Модульная организация сети ограничивала более сложные синтаксические структуры. Выбор и гибкость порядка синтаксических категорий требовали повторения целых модулей группировки слов, что подразумевает избыточность слов и усилия по их обслуживанию.
Самоактивация посредством предварительной активности Из-за линейного суперпозиции предварительной активации, передаваемой подключенными микросхемами, и информации афферентного слова в одной популяции mCMC, агрегирование предварительной активации позволило осуществить самоактивацию mCMC.
Далее мы развиваем эту модель и используем наши выводы о зависимой от состояния обработке и функциональной специализации mCMC в иерархических сетях.Мы разработали распределенную сеть синтаксического анализа с 17 взаимодействующими mCMC в качестве сетевых узлов (см. Рис. 11b), эффективно реализовав метасхемы построения структуры (см. Рис. 11a). Каждая сеть слов содержит один узел слов (узлы 13–17 на рис. 11b), представляющий акустическую словоформу, и от 1 до 3 узлов синтаксиса (узлы 1–12 на рис. 11b) для потенциальных синтаксических ролей слова могу предположить. Например, слово паутина существительного «вор» содержит субъектные и объектные узлы (см. Рис.11б). Другие составляющие сети слов, такие как многочисленные семантические ассоциации, для простоты опущены. В каждой сети слов выходные данные узла слова (Py) проецируются на входы (EIN) узлов синтаксиса. Обратные связи от синтаксических узлов к остальной сети слов (включая словарные узлы), которые необходимы для переназначения синтаксических ролей на фактическое семантическое содержимое, для простоты опущены. Все синтаксические узлы, представляющие конкретную синтаксическую роль (например, субъект) в различных сетях слов, образуют синтаксических пулов , где они связаны взаимно запрещающими связями (опущены на рис.11б для ясности). Это гарантирует, что конкретную роль может взять на себя только одно слово (в то время как слово может выполнять несколько ролей, как в предложении «Я рисую стену на стене»). Наконец, синтаксические пулы связаны согласно синтаксическим правилам (например, субъекты к глаголам, но не к объектам). Если два синтаксических пула подключены, все синтаксические узлы в исходном пуле соединяются в режиме «все для всех» с популяциями Py всех синтаксических узлов в целевом пуле. Проекция на Py вызывает повышение возбудимости (облегчение) без фактической активации узла (таким образом, устанавливая ожидание).Подобно модели Kunze et al. (2017), контекстная информация направляет построение структуры с помощью тормозных связей, чтобы устранить двусмысленность в предложениях.
Рис. 11Архитектура сети синтаксического анализа. a Метасхема построения структуры использовалась в качестве строительного блока для сети синтаксического анализа. Мы интерпретировали верхний mCMC как узел синтаксиса , представляющий синтаксические категории, а нижний как узел слов , представляющий отдельные слова словаря.Обратите внимание, что для каждой роли, которую может принимать слово, может быть несколько узлов синтаксиса. b В сети синтаксического анализа 17 взаимодействующих mCMC представлены либо узлами синтаксиса (1–12), либо узлами слов (13–17). Одно слово и несколько узлов синтаксиса образуют сеть слов (с цветовой кодировкой), в то время как все узлы синтаксиса одного вида образуют синтаксический пул (пунктирные рамки). Узлы Word однонаправленно проецируются на узлы синтаксиса через прямые соединения (черные стрелки). Узлы синтаксиса в разных пулах связаны между собой боковыми связями (серые стрелки) и обмениваются вспомогательными сигналами, которые обусловливают установление устойчивых паттернов активности.Взаимное запрещение внутри синтаксических пулов гарантирует, что конкретную синтаксическую роль принимает только одно слово (не показано для простоты). Контекстная информация помогает семантической интерпретации (пунктирные стрелки)
Для моделирования восприятия предложения узлы слов воспринимали краткие акустические входы и передавали свою временную активацию соответствующим узлам синтаксиса. Среди синтаксических узлов активировались только те, которые были ранее синтаксически предсказаны, т. Е. Получили боковой стимулирующий сигнал.Первоначальное ожидание восприятия предложения моделируется как начальный управляющий сигнал, принимаемый узлом синтаксиса, отражающим субъект. Важные параметры языковой модели приведены в Таблице 3.
Таблица 3 Важные параметры языковой моделиДля моделирования процесса синтаксического анализа мы рассмотрели предложение «Я ударил вора дубинкой» (Kunze et al., 2017) . В процессе синтаксического анализа узлы слов реагировали на их последовательную стимуляцию (верхний график на рис. 12а) и выборочно активировали синтаксические узлы (нижние графики на рис.12а) — обеспечение непрерывного отслеживания активности в конце процесса синтаксического анализа. Таким образом, сохраняется информация об активированных синтаксических ролях (совокупные сигналы каждого синтаксического пула) и фактической сети слов, связанной с этой ролью (какой узел в синтаксическом пуле активируется, обратите внимание, что из-за взаимного запрета только один узел на пул может быть активным).
Рис. 12Анализ синтаксической сети. a Верхний график: узлы слова реагируют на свою последовательную стимуляцию (серые области) и выборочно активируют узлы синтаксиса.Нижний график: активации синтаксических пулов как суммы их соответствующих синтаксических узлов. b Изменение контекстной информации, т. Е. Блокирование модификатора объекта, изменяет интерпретацию предложения
В зависимости от контекстной информации фраза с клубом дополнительно определяет глагол hit , т. Е. Действует как наречная фраза , или объект , вор , т. е. действующий как прилагательное словосочетание (Kunze et al. 2017). Разбор предложения «вор ударил вора дубинкой» также возможен благодаря разделению лексической информации о словах и синтаксических категорий, что позволяет использовать несколько экземпляров слов независимо от их синтаксических категорий.
Особенность метасхемы построения структуры значительно увеличивает достоверность сети синтаксического анализа, поскольку она позволяет рассматривать более сложные синтаксические структуры. Восприятие отдельных слов (например, вор) является временным в узлах слов и возбуждает все связанные синтаксические узлы (например, S, O 1 , O 2 ). Однако возбуждение остается только для синтаксически предсказанных узлов синтаксиса (S), но исчезает для непредсказуемых узлов, которые не принимают вспомогательный сигнал (O 1 , O 2 ).
Введение в нейроны и нейронные сети | Раздел 1, Вступительная глава | Neuroscience Online: Электронный учебник для неврологии | Кафедра нейробиологии и анатомии
Три фунта желеобразного материала, обнаруженные в наших черепах, — это самая сложная машина на Земле и, возможно, во Вселенной. Его феноменальные свойства были бы невозможны без сотен миллиардов нейронов, из которых он состоит, и, что важно, без связей между этими нейронами. К счастью, многое известно о свойствах отдельных нейронов и простых нейронных сетей, и аспекты сложных нейронных сетей начинают разгадываться.Эта глава начнется с обсуждения нейрона, элементарного узла или элемента мозга, а затем перейдет к обсуждению способов, которыми отдельные нейроны общаются друг с другом. Что делает нервную систему таким фантастическим устройством и отличает мозг от других органов тела, так это не то, что в ней есть 100 миллиардов нейронов, а то, что нервные клетки способны взаимодействовать друг с другом таким высокоструктурированным образом, что формируют нейронные сети. . Чтобы понять нейронные сети, необходимо понять способы, которыми один нейрон взаимодействует с другим через синаптические связи, и процесс, называемый синаптической передачей.Синаптическая передача бывает двух основных видов: возбуждение и торможение. Всего несколько взаимосвязанных нейронов (микросхема ) могут выполнять сложные задачи, такие как опосредовать рефлексы, обрабатывать сенсорную информацию, генерировать движения и опосредовать обучение и память. Более сложные сети (макросхемы , ) состоят из множества встроенных микросхем. Макросхемы опосредуют высшие функции мозга, такие как распознавание объектов и познание. Итак, в нервной системе повсеместно встречаются сети нескольких уровней.Сети также преобладают в пределах нейронов. Эти наноцепи составляют базовый биохимический механизм для обеспечения ключевых свойств нейронов, таких как обучение и память, а также генезиса ритмичности нейронов.Нейрон
Основные морфологические особенности нейронов
100 миллиардов нейронов в головном мозге имеют ряд общих черт (рис. 1). Нейроны отличаются от большинства других клеток тела тем, что они поляризованы и имеют различные морфологические области, каждая из которых выполняет определенные функции. Дендриты — это область, где один нейрон получает связи от других нейронов. Тело клетки или сома содержит ядро и другие органеллы, необходимые для клеточной функции. Аксон является ключевым компонентом нервных клеток, по которому информация передается от одной части нейрона (например, тела клетки) к концевым областям нейрона. Аксоны могут быть довольно длинными, достигая метра или около того в некоторых сенсорных и двигательных нервных клетках человека.Синапс является конечной областью аксона, и именно здесь один нейрон образует соединение с другим и передает информацию посредством процесса синаптической передачи . Нейрон цвета морской волны на рисунке 1 (щелкните «Нейрон, связанный с постсинаптическим нейроном») называется постсинаптическим нейроном. Терминал коричневого цвета слева, следовательно, называется пресинаптическим нейроном. Один нейрон может получать контакты от множества разных нейронов.На рисунке 1 (щелкните «Нейрон, получающий синаптический вход») показан пример трех пресинаптических нейронов, контактирующих с одним постсинаптическим нейроном коричневого цвета, но было подсчитано, что один нейрон может получать контакты до 10 000 других клеток. Следовательно, потенциальная сложность сетей огромна. Точно так же любой нейрон может контактировать до 10 000 постсинаптических клеток. (Обратите внимание, что нейрон коричневого цвета, который был пресинаптическим по отношению к нейрону цвета морской волны, является постсинаптическим по отношению к розовым, зеленым и синим нейронам.Таким образом, большинство «пресинаптических» нейронов являются «постсинаптическими» по отношению к некоторым другим нейронам.Рисунок 1 (щелкните «Синапс») также показывает расширенный вид синапса. Обратите внимание, что пресинаптическая клетка не связана напрямую с постсинаптической клеткой. Они разделены промежутком, известным как синаптическая щель . Следовательно, чтобы общаться с постсинаптической клеткой, пресинаптический нейрон должен выпускать химический мессенджер. Этот мессенджер находится в везикулах , содержащих нейротрансмиттер, (синие точки обозначают нейромедиатор).Потенциал действия, который проникает в пресинаптический терминал, заставляет эти везикулы сливаться с внутренней поверхностью пресинаптической мембраны и высвобождать их содержимое посредством процесса, называемого экзоцитозом . Освободившийся передатчик диффундирует через промежуток между пре- и постсинаптической клеткой и очень быстро достигает постсинаптической стороны синапса, где связывается со специализированными рецепторами, которые «распознают» передатчик. Связывание с рецепторами приводит к изменению проницаемости ионных каналов в мембране и, в свою очередь, к изменению мембранного потенциала постсинаптического нейрона, известного как постсинаптический синаптический потенциал (PSP) .Таким образом, передача сигналов между нейронами связана с изменениями электрических свойств нейронов. Чтобы понять нейроны и нейронные цепи, необходимо понимать электрические свойства нервных клеток.
Потенциалы покоя и действия
Потенциалы покоя. На рисунке 2 показан пример идеализированной нервной клетки. Во внеклеточную среду помещен микроэлектрод. Микроэлектрод — это не что иное, как небольшой кусок стеклянной капиллярной трубки, который растягивается под действием тепла, образуя очень тонкий наконечник, порядка 1 микрона в диаметре.Микроэлектрод заполняется проводящим раствором и затем подключается к подходящему записывающему устройству, например, осциллографу или самописцу. Когда электрод находится вне клетки во внеклеточной среде, регистрируется нулевой потенциал, поскольку внеклеточная среда является изопотенциальной. Однако, если электрод проникает в ячейку так, что кончик электрода теперь находится внутри ячейки, на записывающем устройстве наблюдается резкое отклонение. Регистрируется потенциал около -60 милливольт внутри отрицательного по отношению к внешней стороне.Этот потенциал называется потенциалом покоя и остается постоянным в течение неопределенных периодов времени при отсутствии какой-либо стимуляции. Если электрод удален, снова регистрируется нулевой потенциал. Потенциалы покоя — это не просто характеристики нервных клеток; все клетки тела обладают потенциалом покоя. Что отличает нервные клетки от других возбудимых мембран (например, мышечные клетки), так это то, что они способны изменять свой потенциал покоя. В случае нервных клеток для интеграции информации и передачи информации, а в случае мышечных клеток — для производства мышечных сокращений.
Потенциалы действия. На рисунке 3 показан еще один эскиз идеализированного нейрона. Этот нейрон был пронзен одним электродом для измерения потенциала покоя и вторым электродом, называемым стимулирующим электродом. Стимулирующий электрод подключается через переключатель к батарее. Если батарея ориентирована так, что положительный полюс подсоединен к переключателю, замыкание переключателя сделает внутреннюю часть элемента несколько более положительной в зависимости от размера батареи.(Такое уменьшение поляризованного состояния мембраны называется деполяризацией .) На фиг.3 показана анимация, в которой переключатель многократно размыкается и замыкается, и каждый раз, когда он замыкается, в схему подключается батарея большего размера. Первоначально замыкание переключателя вызывает только небольшие деполяризации. Однако потенциалы становятся больше, и в конечном итоге деполяризация становится достаточно большой для запуска потенциала действия , также известного как спайк или импульс .Потенциал действия связан с очень быстрой деполяризацией, позволяющей достичь пикового значения около +40 мВ всего за 0,5 миллисекунды (мс). За пиком следует столь же быстрая фаза реполяризации .
Напряжение, при котором деполяризация становится достаточной для запуска потенциала действия, называется порогом . Если батарея большего размера используется для генерации надпороговой деполяризации, единичный потенциал действия по-прежнему генерируется, и амплитуда этого потенциала действия такая же, как потенциал действия, запускаемый только пороговым стимулом.Простая запись на рисунке 3 иллюстрирует две очень важные особенности потенциалов действия. Во-первых, они вызываются по схеме «все или ничего» . Либо потенциал действия вызывается стимулами на уровне порога или выше, либо потенциал действия не проявляется. Во-вторых, потенциалы действия — это очень короткие события продолжительностью всего несколько миллисекунд. Запуск потенциала действия в некоторой степени аналогичен подаче спички на предохранитель. Определенная температура необходима для воспламенения предохранителя (т.е.е., предохранитель имеет порог). Спичка, выделяющая большее количество тепла, чем пороговая температура, не приведет к тому, что предохранитель будет гореть ярче или быстрее. Так же, как потенциалы действия вызываются по принципу «все или ничего», они также распространяются по принципу «все или ничего». Как только потенциал действия инициируется в одной области нейрона, такой как тело клетки, этот потенциал действия будет распространяться по аксону (как горящий предохранитель) и в конечном итоге проникнет в синапс, где он может инициировать процесс синаптической передачи.
В примере на Рисунке 3 был сгенерирован только один потенциал действия, потому что продолжительность каждого из двух надпороговых стимулов была настолько короткой, что было достаточно времени только для инициирования единственного потенциала действия (т. Е. Стимул закончился раньше, чем потенциал действия завершил свой цикл деполяризации-реполяризации). Но, как показано в анимации на Рисунке 4, более продолжительные стимулы могут привести к инициированию множественных потенциалов действия, частота которых зависит от интенсивности стимула.Таким образом, очевидно, что нервная система кодирует информацию не с точки зрения изменения амплитуды потенциалов действия, а, скорее, с точки зрения их частоты. Это очень универсальное свойство. Чем выше интенсивность механического стимула к сенсорному рецептору, тем больше количество потенциалов действия; чем сильнее растягивается рецептор растяжения мышцы, тем больше количество потенциалов действия; чем больше интенсивность света, тем большее количество потенциалов действия передается в центральную нервную систему.Точно так же в двигательной системе, чем больше количество потенциалов действия в двигательном нейроне, тем сильнее будет сокращение мышцы, которая получает синаптическую связь от этого двигательного нейрона. Инженеры называют этот тип кодирования информации частотно-импульсной модуляцией .
Синаптические потенциалы и синаптическая интеграция
На рисунке 5 показаны три нейрона. Зеленый цвет будет называться возбуждающим нейроном по причинам, которые вскоре станут ясны.Он соединяется с постсинаптическим нейроном синего цвета. Следы ниже (нажмите «Play») иллюстрируют последствия запуска потенциала действия в зеленом нейроне. Этот потенциал действия в пресинаптическом нейроне приводит к снижению мембранного потенциала постсинаптической клетки. Мембранный потенциал изменяется от значения покоя около -60 милливольт до более деполяризованного состояния. Этот потенциал называется возбуждающим постсинаптическим потенциалом (ВПСП) .Он является «возбуждающим», потому что перемещает мембранный потенциал к пороговому значению, и является «постсинаптическим», потому что это потенциал, зарегистрированный на постсинаптической стороне синапса. Обычно (и это важный момент) единичный потенциал действия в пресинаптической клетке не производит ВПСП, достаточно большого, чтобы достичь порога и запустить потенциал действия. Но, если в пресинаптической клетке запускаются множественные потенциалы действия, соответствующие множественные возбуждающие потенциалы могут суммироваться посредством процесса, называемого временным суммированием , для достижения порога и запуска потенциала действия.ВПСП можно рассматривать как «сигнал движения» постсинаптическому нейрону для передачи информации по сетевому пути.
Нейрон, окрашенный в красный цвет на рисунке 5, называется тормозным нейроном. Как и зеленый нейрон, он также устанавливает синаптический контакт с синим постсинаптическим нейроном. Он также высвобождает химический передатчик-мессенджер, но последствия связывания передатчика от синей клетки с рецепторами на постсинаптической клетке противоположны последствиям передатчика, выпущенного зеленым нейроном.Следствием потенциала действия красного пресинаптического нейрона является увеличение мембранного потенциала синего постсинаптического нейрона. Мембранный потенциал более отрицательный, чем был раньше (гиперполяризация ), и поэтому мембранный потенциал дальше от порогового значения. Этот тип потенциала называется ингибирующим постсинаптическим потенциалом (IPSP) , потому что он предотвращает запуск постсинаптического нейрона потенциала действия. Это «стоп-сигнал» для постсинаптической клетки.Таким образом, зеленый нейрон говорит «вперед», а красный нейрон говорит «стоп». Что же делать постсинаптическому нейрону?
Нейроны похожи на счетные машины. Они постоянно суммируют возбуждающий и тормозной синаптический вход во времени (временное суммирование) и по площади дендритов, получающих синаптические контакты ( пространственное суммирование) , и если это суммирование находится на пороговом уровне или выше, они запускают потенциал действия. Если сумма ниже порога, потенциал действия не инициируется.Этот процесс называется синаптической интеграцией и проиллюстрирован на рисунке 5. Первоначально два потенциала действия в зеленом нейроне вырабатывали суммирующие ВПСП, которые запускали потенциал действия в синем нейроне. Но если IPSP от тормозного нейрона возникает непосредственно перед двумя потенциалами действия в возбуждающем нейроне, сумма одного IPSP и двух EPSP ниже порогового значения, и в постсинаптической клетке не возникает никакого потенциала действия. Тормозящий нейрон (и подавление в целом) — это способ стробирования или регулирования способности возбуждающего сигнала запускать постсинаптическую клетку.
Нейронные сети
Мотивы микросетей
Как указывалось ранее в этой главе, нейрон может получать контакты до 10 000 пресинаптических нейронов, и, в свою очередь, любой нейрон может контактировать до 10 000 постсинаптических нейронов. Комбинаторная возможность может привести к чрезвычайно сложным нейронным цепям или сетевым топологиям , которые могут быть очень трудными для понимания. Но, несмотря на потенциально огромную сложность, многое можно узнать о функционировании нейронных цепей, исследуя свойства подмножества простых конфигураций цепей.На рисунке 6 показаны некоторые из этих мотивов микросхем или микросетей. Несмотря на простоту, они могут делать многое из того, что должна делать нервная система.
Возбуждение с прямой связью . Позволяет одному нейрону передавать информацию своему соседу. Длинные цепочки из них могут использоваться для передачи информации через нервную систему.
Ингибирование с прямой связью .Пресинаптическая клетка возбуждает тормозящий интернейрон (интернейрон — это нейрон, расположенный между двумя нейронами), и этот тормозной интернейрон затем подавляет следующую клетку-последователь. Это способ отключения или ограничения возбуждения в нейроне, расположенном ниже по течению в нейронной цепи.
Схождение / расхождение . Одна постсинаптическая клетка получает конвергентный вход от ряда различных пресинаптических клеток, и любой отдельный нейрон может создавать дивергентные связи со многими разными постсинаптическими клетками.Дивергенция позволяет одному нейрону общаться со многими другими нейронами в сети. Конвергенция позволяет нейрону получать входные данные от многих нейронов в сети.
Боковое ингибирование . Пресинаптическая клетка возбуждает тормозящие интернейроны, и они подавляют соседние клетки в сети. Как подробно описано ниже в этой главе, этот тип схемы может использоваться в сенсорных системах для улучшения контуров.
Обратное / повторяющееся подавление .На панели E1 пресинаптическая клетка соединяется с постсинаптической клеткой, а постсинаптическая клетка, в свою очередь, соединяется с интернейроном, который затем подавляет пресинаптическую клетку. Этот контур может ограничивать возбуждение в пути. Некоторое начальное возбуждение будет отключено после того, как красный интернейрон станет активным. На панели E2 каждый нейрон в замкнутой цепи подавляет нейрон, с которым он связан. Казалось бы, эта схема ничего не делает, но, как будет показано далее в этой главе, она может привести к генерации сложных паттернов всплесковой активности.
Обратная связь / рекуррентное возбуждение . На панели F1 пресинаптический нейрон возбуждает постсинаптический нейрон, а этот постсинаптический нейрон возбуждает пресинаптический нейрон. Этот тип цепи может выполнять функцию переключателя, потому что, как только пресинаптическая клетка активируется, эта активация может сохраняться. Активация пресинаптического нейрона могла включить эту сеть, и она могла остаться. На панели F2 показаны варианты возбуждения по обратной связи, при которых пресинаптический нейрон возбуждает постсинаптический нейрон, который может по обратной связи возбуждать сам себя (a, аутапс , ) или другие нейроны, которые в конечном итоге имеют обратную связь (b) с самим собой.
Эти простые мотивы — вездесущие компоненты многих нейронных цепей. Давайте рассмотрим несколько примеров того, что могут делать эти сети.
Возбуждение с прямой связью и подавление с прямой связью
Одна из наиболее изученных микросхем — это схема, которая обеспечивает простое рефлекторное поведение. На рисунке 7 показана схема так называемого рефлекса рефлекса коленного рефлекса или растяжения. Невролог ударяет резиновым молоточком по колену, в результате чего нога разгибается.Этот тест используется как простой способ проверить целостность некоторых сенсорных и моторных путей в спинном мозге. Удар молотка растягивает мышцу и приводит к инициированию потенциалов действия в сенсорных нейронах внутри мышцы, которые чувствительны к растяжению. (Потенциалы действия представлены маленькими яркими «огоньками» на анимации.) Потенциалы действия инициируются по принципу «все или ничего» и распространяются в спинной мозг, где аксон разделяется (раздваивается) на две ветви.
Давайте сначала обсудим ветвь слева, которая формирует синаптическую связь (зеленый треугольник) с Extensor (E) мотонейроном (окрашен в синий цвет). Потенциал действия сенсорного нейрона вторгается в синаптический терминал сенсорного нейрона, вызывая высвобождение медиатора и последующее возбуждение двигательного нейрона. Растяжение мышцы приводит к возникновению потенциала действия в двигательном нейроне (МН), который затем распространяется по периферическому нерву и вторгается в синапс в мышце, вызывая высвобождение передатчика и потенциал действия в мышце.Потенциал действия в мышечной клетке приводит к сокращению мышцы и разгибанию конечности. Итак, здесь у нас есть простая схема возбуждения с прямой связью, которая опосредует поведение.
Теперь давайте рассмотрим правую ветвь аксона сенсорного нейрона на рисунке 7. Потенциал действия сенсорного нейрона вторгается в синаптический терминал сенсорного нейрона, вызывая высвобождение медиатора и последующее возбуждение постсинаптического интернейрона, окрашенного в черный цвет.Этот нейрон называется интернейроном , потому что он расположен между одним нейроном (здесь SN) и другим нейроном (здесь MN). Возбуждение интернейрона приводит к инициированию действия и последующему высвобождению передатчика из пресинаптического терминала интернейрона (черный треугольник), но для этой ветви цепи передатчик ведет к IPSP в постсинаптическом сгибателе (F ) мотонейрон (окрашен в красный цвет). Функциональные последствия этого подавления прямой связи заключаются в уменьшении вероятности того, что двигательный нейрон-сгибатель станет активным и вызовет несоответствующее сгибание ноги.
Схождение и расхождение
Упрощенная схема, опосредующая рефлекс растяжения, представлена на рисунке 8. Однако правильное функционирование схемы рефлекса растяжения также зависит от конвергенции и расхождения. Один сенсор имеет несколько ветвей, которые расходятся и образуют синаптические связи со многими отдельными моторными нейронами (щелкните «Дивергенция»). Следовательно, когда мышца сокращается в результате нажатия невролога, это происходит потому, что несколько мышечных волокон активируются одновременно несколькими двигательными нейронами.Кроме того, когда мышца растягивается, активируется не один, а несколько сенсорных нейронов, и все эти сенсорные нейроны проецируются в спинной мозг, где они сходятся к отдельным моторным нейронам-разгибателям (нажмите «Конвергенция»). Итак, рефлекс растяжения возникает из-за комбинированных эффектов активации нескольких сенсорных нейронов и моторных нейронов-разгибателей.
Боковое торможение
Улучшение кромки.Боковое торможение очень важно для обработки сенсорной информации. Одним из примеров является явление в зрительной системе, называемое усилением краев. На рисунке 9 показаны две полосы: темно-серая полоса слева и светло-серая полоса справа. Хотя темная полоса и светлая полоса имеют одинаковую яркость по всему полю, при внимательном рассмотрении обнаруживается, что светло-серая полоса кажется несколько светлее на границе темно-серой полосы, чем в других областях поля.Напротив, темно-серая полоса на границе выглядит несколько темнее, чем в других областях темного поля. Это явление улучшения границ, которое помогает визуальной системе извлекать важную информацию из визуальных сцен. Усиление краев опосредуется, по крайней мере частично, боковым торможением в сетчатке.
Давайте сначала рассмотрим схему без бокового торможения (Рисунок 10, щелкните «Без бокового торможения»). Свет падает на сетчатку (Часть A), а его интенсивность можно описать ступенчатым градиентом (Часть B).Для упрощения предположим, что темно-серая область имеет интенсивность пяти единиц, а светло-серая область — десять единиц. Градиент света активирует фоторецепторы, и фоторецепторы создают синаптические связи с нейронами второго порядка. Предположим, что интенсивность света 5 единиц приводит к 5 всплескам / с, а интенсивность света 10 единиц приводит к 10 всплескам / с (Часть C) в фоторецепторах, и что синаптическая сила достаточна (здесь обозначена как +1) так что интенсивность света 5 единиц приводит к 5 импульсам / с, а интенсивность света 10 единиц приводит к 10 импульсам / с (Часть C) соответственно в нейронах второго порядка.Если никакой дальнейшей обработки информации не происходит, воспринимаемый градиент будет точно таким же, как градиент интенсивности света (часть B, красный след). Но это не то, что воспринимается, и латеральное торможение объясняет разницу.
Теперь рассмотрим расширенную схему с боковым торможением (нажмите «С боковым торможением»). Каждый из фоторецепторов устанавливает тормозные синаптические связи со своим соседним нейроном второго порядка. Сила подавления (обозначается -0.2) меньше силы возбуждения (пожертвовано +1). Прежде чем смотреть на границу, рассмотрите выход схемы в однородных областях каждого поля. Далеко справа от границы все клетки получают одинаковое возбуждение и одинаковое торможение. Без бокового торможения интенсивность света в 10 единиц произвела бы 10 импульсов в секунду в нейроне второго порядка. Но из-за тормозящей связи от соседних нейронов справа и слева выходная мощность снижается до 6 импульсов в секунду.То же самое верно для клеток, расположенных далеко слева от границы, но величина возбуждения меньше и, соответственно, величина торможения меньше. Ключевая обработка происходит на границе или краю. Обратите внимание, что нейрон справа от границы получает такое же подавление от нейрона справа от нее, но получает меньшее подавление от нейрона слева от него на другой стороне границы. Следовательно, он получает больше чистого возбуждения и имеет выходной сигнал 7 пиков / с, а не 6 пиков / с его соседа справа.Теперь посмотрите на нейрон слева от границы. Он получает слабое торможение от своего соседа слева, но более сильное торможение от своего соседа справа по другую сторону границы. Следовательно, он получает меньше чистого возбуждения и имеет выходную мощность 2 пика / с, а не 3 пика / с его соседа слева. Таким образом, в результате бокового торможения информация, передаваемая нервной системе, и воспринимаемый градиент будут версией исходной с усиленной границей или краем (рис. 10B)!
диапазонов Маха.Простая схема сетчатки с боковым торможением может объяснить феномен усиления краев. Это также может объяснить визуальную иллюзию, известную как полосы Маха. На рисунке 11 показан градиент светлых и темных вертикальных полос, поперек которых проходит тонкая горизонтальная линия. Похоже, что горизонтальная линия имеет неравномерное распределение интенсивности: она темнее в области светлых вертикальных градиентов и светлее в области темных вертикальных градиентов. Это визуальная иллюзия .Иллюзию можно раскрыть, поместив маску поверх вертикального градиента. (Нажмите «Play», чтобы добавить маску.) Теперь вы можете видеть, что горизонтальная полоса имеет равномерную интенсивность. В некоторых областях он воспринимается более темным, потому что клетки сетчатки, которые реагируют на более темную область горизонтальной полосы, сильно подавляются клетками, реагирующими на яркую область вертикальной полосы. Напротив, полоса воспринимается ярче в некоторых областях, потому что клетки сетчатки, которые реагируют на более светлую область горизонтальной полосы, только слабо ингибируются клетками, реагирующими на темную область вертикальной полосы.
Обратная связь / повторное торможение
Запрет обратной связи в микросхемах. Подавление обратной связи играет общую роль в гашении возбуждения через нейронную цепь. Классическим примером является клетка Renshaw в спинном мозге. Аксон ветви спинномозгового мотонейрона. Одна ветвь иннервирует мышцу, как описано ранее (например, рис. 7), а другая ветвь создает возбуждающую синаптическую связь с интернейроном, называемым клеткой Реншоу.Интернейрон, в свою очередь, подавляет двигательный нейрон, замыкая петлю. Другой пример подавления обратной связи обнаружен в гиппокампе. Пирамидные клетки СА3-типа создают возбуждающие связи с клетками корзины и обратную связь с клетками корзины, чтобы ингибировать клетки СА3. Термин повторяющееся ингибирование применяется к простым схемам подавления обратной связи, таким как цепь Реншоу в спинном мозге и цепь корзиночных клеток в гиппокампе.
Подавление обратной связи в наносхемах.Подавление обратной связи распространено не только во многих нейронных цепях; он также широко распространен в биохимических цепях. Здесь он может служить подложкой для генерации колебаний. Они могут охватывать несколько временных масштабов от секунд до дней в зависимости от молекулярных компонентов схемы.
Рис. 12. Данные Byrne, Canavier, Lechner, Clark and Baxter, 1996.
- Эндогенное взрывное поведение в нейронах.Идеализированный нейрон, описанный ранее в этой главе, молчал в отсутствие стимуляции (например, рисунок 3). Однако некоторые нейроны активируют потенциалы действия в отсутствие стимуляции, и в некоторых случаях паттерны возбуждения могут демонстрировать взрывной паттерн, в котором последовательные высокочастотные всплески активности сменяются периодами покоя. Такие нейронные свойства могут быть важны для создания ритмического поведения, такого как дыхание. На рисунке 12 показан пример записи нейрона беспозвоночного, имеющего эндогенный импульсный ритм.Этот конкретный нейрон называется параболическим всплеском, потому что характер интервалов между всплесками является длинным в начале и в конце цикла всплеска, но очень коротким в середине цикла. Клетка запускает всплеск потенциалов действия, а затем замолкает, но вскоре происходит еще один всплеск, и этот процесс продолжается бесконечно каждые десять-пятнадцать секунд. Взрыв происходит, даже если нейрон хирургическим путем удален из ганглия и помещен в культуру, чтобы не было синаптических связей с другими нейронами.Таким образом, для этого ритма не требуется нейронная сеть — она эндогенная. Но это действительно связано с наноконтролем внутри клетки. На рисунке 13A показана очень упрощенная версия этой сети, в которой подчеркивается ключевой принцип работы. Критическим для этой сетевой функции является канал в
мембрана (обозначена g SI ), проницаемая для Ca 2+ . Поскольку концентрация Ca 2+ относительно высока во внеклеточной среде и низка внутри клетки, Ca 2+ будет двигаться вниз по градиенту концентрации и тем самым деполяризует клетку.В конце концов, деполяризация достигает порога, и клетка начинает срабатывать. Обжиг приводит к дополнительному притоку Ca 2+ (зеленая стрелка) и накоплению Ca 2+ внутри клетки. Ключевым этапом является то, что накопление Ca 2+ ингибирует (красная стрелка) дальнейший приток кальция и прекращает всплеск. Всплеск остается прекращенным, пока уровни внутриклеточного Ca 2+ остаются повышенными. Но уровни Ca 2+ не остаются повышенными надолго.Они восстанавливаются внутриклеточными буферами и удаляются из клетки с помощью насосов (I NaCa и I CaP ). Когда внутриклеточные уровни Ca 2+ снижаются, ингибирование канала снимается (растормаживание , ), и нейрон снова начинает деполяризоваться, и начинается новый всплеск. По сути, то, что мы видим здесь на уровне наносети, является повторением сети, препятствующей обратной связи (рис. 13B). Первоначальный процесс возбуждения приводит к активации тормозного процесса, который отключается от него и прекращает его.В такой сети возникнут колебания, если возбуждающее побуждение будет непрерывным, но тормозящий процесс ослабевает по своей эффективности.
Циркадные ритмы.Второй пример подавления обратной связи — это наноцепь регуляции генов, лежащих в основе циркадных ритмов. Циркадный ритм позвоночных обусловлен работой группы нейронов в области мозга, называемой супрахиазматическим ядром, которая расположена чуть выше зрительного нерва. Эти нейроны оказывают сильное влияние как на высвобождение гормонов, таких как мелатонин, кортизол, так и на вегетативные функции, такие как температура тела (рис. 14). Несмотря на огромное влияние этого генератора, его работа сводится к очень простой схеме, и действительно, не к нейронной схеме, а к другой наносхеме.Основной механизм, по-видимому, сохраняется у всех видов животных, включая человека. На рисунке 15 представлена упрощенная принципиальная схема основных компонентов. Участвует несколько генов, но основной механизм включает ген под названием на , где на — на период. Этот ген был впервые идентифицирован у плодовой мухи Drosophila , но также присутствует у позвоночных. Ген на приводит к продукции на матричной РНК. на мРНК покидает ядро и попадает в цитоплазму, где приводит к синтезу белка PER.PER диффундирует или транспортируется обратно в ядро, где репрессирует дальнейшую транскрипцию гена на . Концептуально эта система очень похожа на механизм разрывного нейрона, рассмотренный выше. Ген активируется, он производит сообщение и белок, а белковая обратная связь подавляет экспрессию гена. Но как цикл повторяется? Ключевым механизмом является деградация PER. Белок PER разлагается и разлагается в течение 24 часов.Так как белок PER разрушается, подавление или подавление снимается (растормаживание), позволяя этому гену снова начать создавать информационную РНК и белок. Итак, как только этот цикл начинается, он повторяется снова и снова через 24 часа. Это основной механизм, лежащий в основе циркадных ритмов и мощного воздействия, которое они оказывают на ряд различных физиологических систем. По сути, все наши циркадные ритмы начинаются с сети подавления молекулярной обратной связи.Рисунок 14. По данным Hastings et al., Nature Rev. Neurosci. , 2003.
Запрет обратной связи в кольцевых цепях.Повторяющееся торможение может, по крайней мере в принципе, объяснить генерацию сложных двигательных паттернов, примером которых является передвижение на четвероногих ногах. Четвероногие животные интересны тем, что четвероногие способны не только двигать своими четырьмя ногами, но и генерировать различные типы циклов активности, называемые походками. На рисунке 16 показаны четыре аллюра. Первая панель — это прогулка (тап по собаке). Последовательность начинается с разгибания левой передней конечности. Затем следуют разгибания правой задней конечности, правой передней конечности и левой задней конечности.На рыси (вторая панель рисунка 16) (постучите по собаке) левая передняя и правая задние конечности находятся в фазе друг с другом и на 180 градусов не совпадают по фазе с правой передней и левой задней конечностями. В связке (третья панель) (постукивание по собаке) левая передняя и правая передние конечности находятся в фазе, но на 180 градусов не совпадают по фазе с левой задней и задней задними конечностями. Галоп (четвертая панель) (постукивание по собаке) — вариант скачек, в котором есть небольшая разница фаз между правой и левой передними и задними конечностями.
Как нервная система генерирует эти походки? И нужны ли для каждой отдельные нейронные цепи? К сожалению, нейробиологи не знают ответов на эти вопросы, но поучительно изучить некоторые возможности. Это подход в области нейробиологии под названием Вычислительная и теоретическая нейробиология . Один из способов создания походки проиллюстрирован на рисунке 17. Возьмите четыре отдельных нейрона, каждый с эндогенной импульсной активностью, подобной тому, который показан ранее на рисунке 12, и назначьте активность в каждом из этих нейронов контролю конкретной конечности.Нейроны могут быть «запущены» так, чтобы у них были соответствующие фазовые соотношения для генерации походки, такие как граница, показанная на рисунке 16. Трудность будет заключаться в том, чтобы запустить нейроны точно в точное время. Другой проблемой могут быть небольшие «дрейфы» в колебательных периодах четырех независимых нейронов, которые со временем приведут к несогласованности паттерна (рис. 18). Эта собака не выиграет ни одной гонки и, вероятно, не сможет ходить.
Итак, очевидно, что нейроны должны быть связаны.Один из способов сделать это — использовать повторяющуюся цепь подавления, состоящую из четырех связанных нейронов, чтобы сформировать так называемую «кольцевую» схему, где каждый нейрон в цепи имеет эндогенную импульсную активность, и каждый нейрон связан с другим с помощью тормозной синаптической связи ( Рисунок 19A).
Рисунок 19. По материалам Canavier, Butera, Dror, Baxter, Clark and Byrne, 1997.
Чтобы получить правильные фазовые соотношения для походок, вместо того, чтобы назначать нейрон 3 правую переднюю конечность, он назначается для управления правой задней конечностью, а нейрон 4 назначается для управления правой передней конечностью (a простой поворот схемы) (Рисунок 19Б).При реализации с помощью компьютерного моделирования эта единственная схема способна генерировать четвероногие ворота. Более того, одна и та же схема с небольшими изменениями свойств отдельных нейронов может генерировать каждую из четырех походок, показанных на рисунке 17 (рисунок 20).
Этот результат указывает на важный момент в отношении нейронных сетей.Чтобы понять их, необходимо понимать не только топологию сети, но и природу связей между нейронами (являются ли они возбуждающими или тормозящими), а также свойства отдельных узлов (т. Е. нейроны). Кроме того, это моделирование иллюстрирует явление, называемое динамической реконфигурацией . Для генерации этих четырех разных походок не обязательно иметь четыре разные сети — все это можно сделать с помощью одной схемы.Фактическая схема, генерирующая четырехпедальную походку, более сложна, чем схема на рис. 19. Заинтересованный читатель может отсылать к недавнему обзору Оле Кейна (см. Дополнительную литературу).
Обратная связь / рекуррентное возбуждение
Периодическое возбуждение в наноцепях и микросхемах имеет решающее значение для процессов обучения и памяти. Обучение включает в себя изменения биофизических свойств нейронов и изменения синаптической силы. Накапливающиеся данные указывают на то, что положительная обратная связь внутри биохимических каскадов и генных сетей является важным компонентом для индукции и поддержания этих изменений.Более того, периодическое возбуждение обнаруживается по крайней мере в некоторых микросхемах, участвующих в процессах памяти. Яркий пример находится в области CA3 гиппокампа.
Рисунок 21. Изменено из Byrne and Roberts, 2009.
На рисунке 21 показаны основные характеристики рекуррентной возбуждающей цепи CA3. Шесть различных пирамидных нейронов гиппокампа обозначены как U , V , W , X , Y и Z .Каждый из этих нейронов получает синаптическую связь от пресинаптических нейронов, обозначенных a , b , c , d , e и f. Эти пресинаптические нейроны могут быть активными или неактивными с 0 и черным цветом, представляющим неактивный нейрон, и 1 и зеленым цветом, указывающим активный. Важным аспектом этой схемы является то, что синаптические связи от входного пути достаточно сильны, чтобы активировать (активировать) пирамидный нейрон, с которым они связаны.Например, если нейрон a активирован, будет активирован нейрон Z , который представлен как 1 на панели вывода. Эта топология — не что иное, как возбуждение с прямой связью. Взаимное возбуждение делает эту схему особенной. Например, нейрон Z и другие пирамидные нейроны имеют коллатерали аксонов, которые посредством обратной связи соединяются друг с другом. Но они не только устанавливают связь между собой. Каждый нейрон устанавливает связь с каждым из четырех других пирамидных нейронов в цепи.Таким образом, каждая пирамида получает сходящуюся информацию от всех других клеток в сети, и, в свою очередь, выход каждого пирамидального нейрона расходится, чтобы установить синаптические связи со всеми другими пирамидными нейронами в цепи. (Следовательно, этот повторяющийся мотив возбуждения вложил в себя мотивы конвергенции и расхождения.) Матрица связности состоит из 36 элементов.
Чтобы эта сеть могла чему-либо научиться, в схему должно быть встроено правило обучения синаптической пластичности.Один из широко распространенных правил известен как Правило обучения Хебба . По сути, он утверждает, что синапс изменит свою силу, если этот синапс активен (то есть высвобождает передатчик) и в то же время активна постсинаптическая клетка. Комбинация этого правила обучения и повторяющегося возбуждающего контура приводит к некоторым интересным эмерджентным свойствам. Например, если нейрон Z активируется входом a, , сила его соединения с самим собой (синапс 1 ) изменится, как показано зеленым синапсом на рисунке 21 (анимация находится в 2-секундном цикле, чтобы иллюстрируют контрольные и модифицированные синапсы).Однако синапс 1 не будет единственным синапсом, который будет усилен. Например, синапс 13 также будет усилен, потому что нейрон Z был активен в то же время, когда нейрон X был активирован вводом c . Напротив, синапс 7 не усиливается, потому что нейрон Y не был активен в то же время, что и нейрон Z . Чистый эффект этой конвергенции и расхождения и правила обучения состоит в том, что исходный входной шаблон активности будет сохранен как изменения в элементах матрицы связности.Поэтому эта схема была названа сетью с автоассоциацией . Важная концепция здесь заключается в том, что «память» находится не в каком-то одном синапсе; это распределено в сети .
Сводка
Значительный прогресс был достигнут в понимании того, как разные простые нейронные сети участвуют в обработке информации и посредническом поведении. Возбуждение с прямой связью и торможение с прямой связью опосредуют рефлекторное поведение.Боковое торможение важно для усиления краев. Повторяющееся возбуждение — важный механизм памяти. Повторяющееся торможение может иметь важное значение для формирования локомоторного поведения. В этих микросхемах заложены сходимость и расхождение. Одни и те же типы сетевых мотивов воспроизводятся в биохимических и генных сетях.
Следующий уровень понимания находится на уровне нейронных сетей, которые опосредуют более сложные, так называемые функции мозга высшего порядка.Их понимание становится возможным благодаря использованию электрофизиологических и оптических методов записи и современных методов визуализации, таких как функциональная магнитно-резонансная томография (фМРТ) и диффузионно-тензорная визуализация (DTI) . ФМРТ позволяет исследователям идентифицировать области мозга, которые заняты когнитивными задачами, тогда как DTI позволяет визуализировать пути, связывающие одну область мозга с другой. Рисунок 22 (любезно предоставлено Тимом Элмором, доктором философии, отдел.из нейрохирургии, Медицинская школа Техасского университета в Хьюстоне) — вид сбоку человеческого мозга, показывающий пути, соединяющие корковые области, выявленные с помощью DTI !.
Рисунок 22. Предоставлено Тимом Элмором, доктором философии.
Распознавание объектов — это пример прогресса в понимании макросхем.Как показано на рисунке 23, обработка визуальной информации начинается в сетчатке, а затем затрагивает несколько областей коры, таких как затылочная кора и височная кора. Внутри этой макросхемы находятся модули, извлекающие информацию более высокого порядка. Предположительно, каждый модуль включает в себя сотни, если не тысячи отдельных микросхем. Задача на будущее — определить, как эти модули работают и как они взаимодействуют с другими модулями. Хотя соединения с прямой связью присутствуют, соединения с обратной связью и боковые соединения широко распространены.Задача огромна, но, возможно, достижению цели будет способствовать использование преимуществ того, что было изучено о принципах работы наносхем и микросхем. Чтобы понять макросхемы, необходимо знать больше, чем топологию сетевых соединений. Необходимо будет знать, как работает каждый модуль, и о динамике межмодульных соединений.
Рисунок 23.Из Феллемана и Ван Эссена, 1991.
Проверьте свои знания
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью
B. Ингибирование с прямой связью
C. Подавление обратной связи
D. Боковое торможение
E.Дивергенция
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью. Этот ответ НЕПРАВИЛЬНЫЙ.
B. Ингибирование с прямой связью
C. Подавление обратной связи
D. Боковое торможение
E. Расхождение
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
А.Прямое возбуждение
B. Запрещение прямой связи. Этот ответ НЕПРАВИЛЬНЫЙ.
C. Подавление обратной связи
D. Боковое торможение
E. Расхождение
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью
B. Ингибирование с прямой связью
С.Подавление обратной связи. Этот ответ НЕПРАВИЛЬНЫЙ.
D. Боковое торможение
E. Расхождение
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью
B. Ингибирование с прямой связью
C. Подавление обратной связи
D. Боковое торможение. Ответ ПРАВИЛЬНЫЙ!
E.Дивергенция
Общие мотивы контуров, которые опосредуют рефлекс растяжения позвоночника, включают все следующие, ЗА ИСКЛЮЧЕНИЕМ:
A. Возбуждение с прямой связью
B. Ингибирование с прямой связью
C. Подавление обратной связи
D. Боковое торможение
E. Расхождение. Этот ответ НЕПРАВИЛЬНЫЙ.
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
А.Пластичность
B. Интеграция
C. Конвергенция
D. Частотно-импульсная модуляция
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
A. Пластичность. Этот ответ НЕПРАВИЛЬНЫЙ.
B. Интеграция
C. Конвергенция
Д.Частотно-импульсная модуляция
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
А. Пластичность
B. Интеграция. Этот ответ ПРАВИЛЬНЫЙ!
C. Конвергенция
D. Частотно-импульсная модуляция
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
А.Пластичность
B. Интеграция
C. Конвергенция Этот ответ НЕПРАВИЛЬНЫЙ.
D. Частотно-импульсная модуляция
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
А. Пластичность
B. Интеграция
C. Конвергенция
Д.Частотно-импульсная модуляция. Этот ответ НЕПРАВИЛЬНЫЙ.
E. Растормаживание
Процесс, посредством которого нейрон суммирует синаптическое возбуждение и торможение, называется:
А. Пластичность
B. Интеграция
C. Конвергенция
D. Частотно-импульсная модуляция
E. Растормаживание Этот ответ НЕПРАВИЛЬНЫЙ.
Сетевой мотив, лежащий в основе циркадных ритмов:
А.Временное суммирование
Б. Пространственное суммирование
C. Подавление обратной связи
D. Возбуждение обратной связи
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
A. Временное суммирование. Этот ответ НЕПРАВИЛЬНЫЙ.
Б. Пространственное суммирование
C. Подавление обратной связи
Д.Возбуждение обратной связи
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
A. Временное суммирование
B. Пространственное суммирование. Этот ответ НЕПРАВИЛЬНЫЙ.
C. Подавление обратной связи
D. Возбуждение обратной связи
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
А.Временное суммирование
Б. Пространственное суммирование
C. Запрет обратной связи. Ответ ПРАВИЛЬНЫЙ!
D. Возбуждение обратной связи
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
A. Временное суммирование
Б. Пространственное суммирование
C. Подавление обратной связи
Д.Возбуждение обратной связи. Этот ответ НЕПРАВИЛЬНЫЙ.
E. Боковое торможение
Сетевой мотив, лежащий в основе циркадных ритмов:
A. Временное суммирование
Б. Пространственное суммирование
C. Подавление обратной связи
D. Возбуждение обратной связи
E. Боковое торможение. Этот ответ НЕПРАВИЛЬНЫЙ.
Автоматическая ассоциация сети важна для:
А.Рефлексы
B. Улучшение кромки
C. Память
D. Передвижение
E. Колебания нейронов
Автоматическая ассоциация сети важна для:
A. Рефлексы. Этот ответ НЕПРАВИЛЬНЫЙ.
B. Улучшение кромки
C. Память
D. Передвижение
E. Колебания нейронов
Автоматическая ассоциация сети важна для:
А.Рефлексы
B. Улучшение края Этот ответ НЕПРАВИЛЬНЫЙ.
C. Память
D. Передвижение
E. Колебания нейронов
Автоматическая ассоциация сети важна для:
А. Рефлексы
B. Улучшение кромки
C. Память Ответ ПРАВИЛЬНЫЙ!
D. Передвижение
E.Колебания нейронов
Автоматическая ассоциация сети важна для:
А. Рефлексы
B. Улучшение кромки
C. Память
D. Передвижение. Этот ответ НЕПРАВИЛЬНЫЙ.
E. Колебания нейронов
Автоматическая ассоциация сети важна для:
А. Рефлексы
Б.Улучшение кромок
C. Память
D. Передвижение
E. Нейрональные колебания. Этот ответ НЕПРАВИЛЬНЫЙ.
Пожертвования Neuroscience Online помогут финансировать разработку новых функций и контента.
Разбираемся в принципе работы К176ИА4.Индикатор цифровой на микросхеме К176II4 K176II4 Описание
Рассматриваемая серия микросхем включает большое количество различных типов счетчиков, большинство из которых работают с кодами веса.
Микросхема К176ИА1 (рис. 172) — шестизначный двоичный счетчик, работающий в коде 1-2-4-8-16-32. Микросхема имеет два входа: вход R устанавливает триггеры на 0, а вход C является входом для подачи счетных импульсов. Установка в 0 происходит при отправке журнала.1 на входе R, переключение триггеров микросхемы — по затуханию импульсов положительной полярности, подаваемых на вход С. при построении
Многовариантные делители частоты Входы с микросхемами должны быть подключены к выходам 32 из предыдущих.Микросхема К176ИА2 (рис. 173) — пятисторонний счетчик, который может работать как двоичный в коде 1-2-4-8-16 при отправке журнала. 1 на управляющем входе A, или как декада с триггером, подключенным к концу декады.0 На входе A. Во втором случае код счетчика 1-2-4-8-10, общий коэффициент деления — 20. Вход R используется для установки триггеров счетчика на 0 подачу в этот журнал. 1. Первые четыре триггера счетчика могут быть переведены в одно состояние подачи журнала. 1 на входах Si — S8. Входы S1 — S8 преобладают над входом R.
Микросхема К176И2 встречается двух разновидностей. Микросхемы ранних выпусков имеют входы cf и cn для подачи тактовых импульсов положительной и отрицательной полярности, соответственно, включенные или.При приложении к входным импульсам CP положительная полярность на входе CN должна быть логарифмической. 1, при подаче на вход CN импульсов отрицательной полярности должен быть лог. 0. В обоих случаях счетчик отключает импульсные отложения.
Другой тип имеет два равноправных входа для подачи тактовых импульсов (выводы 2 и 3), собираемых I. Учет происходит по спаду импульсов положительной полярности, подаваемых на любой из этих входов, и лог необходимо подавать. ко второму из этих входов.1. Можно подавать импульсы и на комбинированные выводы 2 и 3. Изученные автором микросхемы, выпущенные в феврале и ноябре 1981 г., относятся к первым видам, выпущенным в июне 1982 г. и июне 1983 г., ко вторым.
Если вывод 3 микросхемы К176ИА2 Подать лог. 1, обе разновидности микросхем на входе КП (вывод 2) работают одинаково.
С лог. 0 На входе и порядок срабатывания триггера, соответствует временной диаграмме, показанной на рис. 174. В этом режиме на выходе P представляет собой выход элемента, а не, входы которого подключены к выходам 1 и 8 измерителя выделяются импульсы отрицательной полярности, фронты которых совпадают с спадом каждого девятого входного импульса, спад — с спадом каждого десятого.
При подключении к микросхеме К176ИА2 в многоразрядном счетчике входы последующих микросхем должны быть подключены к выходам 8 или 16/10 напрямую, для входа на входы CN. 1. В момент включения напряжения питания триггеры микросхемы К176и могут быть установлены в произвольное состояние. Если при этом счетчик включен в режим десятичного времени, то есть на входе А ведется журнал. 0, а это состояние больше 11, счетчик «зацикливается» между состояниями 12-13 или 14-15.В этом случае на выходах 1 и p формируются им-импульсы с частотой, в 2 раза меньшей частоты входного сигнала. Для выхода из такого режима необходимо установить счетчик в нулевое состояние подачи импульсов на вход R. Обеспечить надежную работу счетчика в десятичном режиме можно, подключив вход и с выходом 4. Тогда, при его значении 12 и более счетчик переходит в бинарный режим Счета и выходит из «запретной зоны», установив после состояния 15 ноль. В моменты перехода из состояния 9 в состояние 10 на вход и с выхода 4 идет лог.0 и счетчик сбрасывается, работает в режиме десятичного счета.
Для индикации состояния декады с помощью микросхемы К176и2 могут использоваться газоразрядные индикаторы, управляемые через децифу К155ИД. Для соответствия микросхеме C155ID1 и K176IA2 можно использовать микросхемы К176ПУ-3 или К561ПУ4 (рис. 175, а) или транзисторы П-Н-П (рис. 175, б).Микросхемы К176ИА3 (рис. 176), К176ИА4 (рис. 177) и К176И5 разработаны специально для использования в электронных часах с семисегментными индикаторами. Микросхема К176ИА4 (рис.177) -декада с преобразователем кода счетчика в код индикатора семерки. Чип имеет три входа — вход R, установка счетчика триггеров в 0 происходит при подаче журнала. 1 На этом входе вход C — переключение триггеров происходит по спаду импульсов положительной полярности
на этом входе. Сигнал на входе S регулирует полярность выходных сигналов.На выходах A, B, C, D, E, F, G — выходные сигналы, выдают числа на семиступенчатом индикаторе, соответствующие состоянию счетчика.При отправке журнала. 0 в журнал управления журналом. 1 На выходах A, B, C, D, E, F, G соответствуют включению соответствующего сегмента. Если вы отправите журнал на input. 1 включение сегментов будет соответствовать бревну. 0 На выходах A, b, C, D, E, F, G. Возможность переключения полярности выходных сигналов значительно расширяет область применения микросхемы.
EXIT P цыплят — перевалочный вывод. Спад импульса положительной полярности на этом выходе формируется в момент перехода счетчика из состояния 9 в состояние 0.
Следует учитывать, что расположение выводов A, B, C, D, E, F, G в паспорте микросхемы и в некоторых справочниках дано для нестандартного расположения сегментов индикатора. На рис. 176, 177 Дана выводы для стандартного расположения сегментов показаны на рис. 111.
Два варианта подключения к микросхеме вакуумных семи индикаторов К176ИА4 с помощью транзисторов показаны на рис. 178. Напряжение UH выбрано в В соответствии с типом используемого индикатора выбор напряжения +25… 30 В на рис. 178 (а) и -15 … 20 В на рис. 178 (b) В некоторых пределах отрегулируйте яркость свечения сегментов индикатора. Транзисторы на схеме. 178 (6) может быть любым кремниевым РНП с обратным током коллекторного перехода не более 1 мкА при напряжении 25 В, если обратный ток транзисторов больше указанного значения или используются транзисторы Германии, между ними аноды и один из выводов индикатора накала Необходимо включить резисторы 30… 60 ком.
Для согласования микросхемы K176IA4 с вакуумными индикаторами удобно использовать микросхемы K168ct2B или K168ct2B (рис. 179), а также KR168ct2B.V, K190T1, K190T2, K161KN1, K161KN2. Подключение микросхемы K161KN1 и K161KN2 показано на рис. 180. При использовании инвертирующей микросхемы K161KN1 на входе S цыпленка K176IA4 протокол должен быть представлен. 1, при использовании несоответствующей микросхемы К161КН2 — лог. 0.
На рис. 181 показаны варианты подключения к микросхеме полупроводниковых индикаторов К176ИА4, на рис.181 (а) с общим катодом, на Рис. 181 (б) — с общим анодом. Резисторы R1 — R7 Требуемый ток устанавливается через сегменты индикатора.Самые маленькие индикаторы могут быть подключены к выходам микросхемы напрямую (рис. 181, Б). Однако из-за большого разброса тока короткого замыкания микросхем, не нормированных техническими условиями, яркость свечения индикаторов также может иметь большой разброс. Частично это можно компенсировать подбором напряжения индикаторов.
Для согласования микросхемы К176ИА4 с полупроводниковыми индикаторами с общим анодом могут использоваться К176ПУ1, К176ПУ2, К176ПУ-3, К561ПУ4, КР1561ПУР4, К561ЛН2 (рис.182). При использовании неконвертирующих микросхем на вход S микросхем должен подаваться лог. 1, при использовании инвертирования — лог. 0.
Согласно схеме Рис. 181 (b), исключив резисторы R1 — R7, можно подключить и немного индикаторы, при этом напряжение питания индикаторов должно быть установлено примерно на 1 с больше номинала, чтобы компенсировать падение напряжения на транзисторах, это напряжение может быть как постоянным, так и пульсирующим, возникающим в результате выпрямления без фильтрации.Жидкокристаллические индикаторы не требуют специального согласования, но для их включения необходим источник прямоугольных импульсов с частотой 30 100 Гц и пластичностью 2, амплитуда импульсов должна соответствовать питанию микросхемы.
Импульсы подаются одновременно на вход микросхемы и на общий электрод индикатора (рис.183) в результате на сегментах, которые необходимо указать, относительно общего электрода индикатора, напряжение с изменяющейся полярностью, на сегментах, которые не следует указывать, напряжение относительно всего электрода равно нулюМикросхема К176и-3 (рис.176) отличается от К176ИА4 тем, что его счетчик имеет коэффициент пересчета 6, а выходной переключатель 1 появляется, когда счетчик установлен в состояние 2.
Микросхема К176ИА5 Содержит кварцевый генератор с внешним резонатором на 32768 Гц и подключенный к нему с Это девятиразрядный делитель частоты и шестизначный делитель частоты, структура микросхемы показана на рисунке 184 (а) типичная схема включения микросхемы показана на рисунке 184 (б) кварцевый кварц подключается. Резонатор, резисторы R1 и R2, КОНДЕНСАТОРЫ C1 и C2 Выходным сигналом кварцевого генератора можно управлять на выходах к и сигнал R с частотой 32768 Гц поступает на вход девятиразрядного двоичного делителя частоты, с его выхода 9 Сигнал с частотой 64 Гц может подаваться на вход 10 шестого делителя, на выходе 14 пятого разряда этого делителя формируется частота 2 Гц, на выходе 15 шестого разряда — 1 Гц.Сигнал с частотой 64 Гц может быть использован для подключения жидкокристаллических индикаторов к выходам микросхем К176И и К176II4.
Вход R служит для сброса триггеров второго делителя и установки фазы колебаний на внешней стороне микросхемы. При обслуживании
Лог. 1 на входе R на выходах 14 и 15 — лог. 0, после удаления лога. 1 На этих выходах появляются импульсы с соответствующей частотой, уменьшение количества импульсов на выходе 15 происходит через 1 секунду после удаления журнала.один.При отправке журнала. 1 Вход S настроен на установку всех триггеров второго делителя в состояние 1 после удаления журнала. 1 От этого входа спад первого импульса на выходах 14 и 15 происходит практически сразу. Обычно вход S постоянно подключен к общему проводу.
Конденсаторы С1 и С2 служат для точной настройки частоты кварцевого генератора. Емкость первого из них может быть в диапазоне единиц до ста пикопарад, емкость второго — -0… 100 пф. С увеличением конденсаторов частота генерации снижается. Точную установку частоты удобнее производить с помощью подстроечных конденсаторов, подключенных параллельно C1 и C2. В этом случае конденсатор, подключенный параллельно С2, осуществляется с грубой настройкой, подключенный параллельно С1 — точной.
Сопротивление резистора R 1 может быть в пределах 4,7 … 68 МОм, но при его значении менее 10 МОм возбуждается
Не все кварцевые резонаторы.Микросхемы К176ИА8 и К561И8-десятичные счетчики с декодером (рис.185). Микросхемы имеют три входа — вход установки исходного состояния R, вход для подачи счетных импульсов отрицательной полярности CN и вход для подачи счетных импульсов положительной полярности CP. Установка счетчика в 0 происходит, когда журнал применяется к входу. 1, а на выходе 0 появляется лог. 1, на выходах 1-9 — лог. 0.
Переключение счетчика происходит по спадам импульсов отрицательной полярности, подаваемых на вход CN, при этом на входе CP должен быть лог.0. Также на вход CP можно подавать импульсы положительной полярности, переключение будет происходить по спаду. При входе в CN должен быть лог. 1. Временная диаграмма микросхемы представлена на рис. 186.Микросхема К561И9 (рис. 187) — счетчик с декодером, работа микросхемы аналогична работе микросхемы К561И8
и К176ИА8, но коэффициент пересчета и количество выходов декодера 8, а не 10. Временная диаграмма микросхемы представлена на рис.188. Также, как и микросхема К561И8, микросхема:К561И9 построена на основе сдвигового регистра с поперечными связями. При подаче напряжения питания и отсутствии разрядного импульса. Триггеры этих микросхем могут находиться в произвольном состоянии, не соответствующем разрешенному состоянию счетчика. Однако в этих микросхемах есть особая цепочка формирования разрешенного состояния счетчика, и при подаче тактичных импульсов счетчик перейдет в нормальный режим работы через несколько тактов.Поэтому в делителях частоты, в которых точная фаза выходного сигнала не важна, на входы R микросхем К176И8, К561И8 и К561И9 допустимо не подавать импульсы начальной установки.
Микросхемы K176I8, K561IA8, K561I9 могут быть объединены в многоразрядные счетчики с последовательной передачей, соединяя выход перехода предыдущей микросхемы с подсекцией CN последующей и подачей на вход Log. 0. Также есть возможность соединить более старый
Выход декодера (7 или 9) с входом следующей микросхемы и подачей в журнал CN.1. Такие способы подключения приводят к появлению задержек в многоразрядном счетчике. Если необходимо, чтобы микросхемы микросхемы многоразрядного счетчика можно было менять одновременно, параллельного переноса с введением дополнительных элементов и нет. На рис. 189 показана схема тройного счетчика с параллельным переносом. Инвертор DD1.1 нужен только для компенсации задержек в элементах DD1.2 и DD1.3. Если не требуется высокая точность одновременности переключения декады счетчика, то входные счетные импульсы могут подаваться на вход CP микросхемы DD2 без инвертора, а на вход CN DD2 — Log.1. Максимальная рабочая частота многоразрядных счетчиков, как с последовательным, так и с параллельным переносом, не снижается относительно частоты отдельной микросхемы.На рис. 190 показан фрагмент схемы таймера с использованием микросхем K176II8 или K561I8. В момент начала ввода микросхемы CN DD1 запускаются счетные импульсы. Когда микросхемы счетчиков установлены в положения, отмеченные на переключателях, журнал появится на всех входах элемента, а не на DD3. 1, элемент
DD3 включится, войдите на выход инвертора DD4.1, сигнализирующий об окончании временного интервала.Микросхемы К561И8 и К561ИА9 Удобно использовать в делителях частоты с переключаемым коэффициентом деления. На рис. 191 показан пример трехкратного делителя частоты. Переключатель SA1 устанавливает единицы необходимого коэффициента пересчета, переключатель SA2 — десятки, переключатель SA3 — сотни. Когда DD1 — DD3 достигает состояния, соответствующего положениям переключателей, журнал поступает на все входы элемента DD4.1. 1.Этот элемент включается и устанавливает триггер на элементах DD4.2 и DD4.3 в состояние, при котором на выходе элемента DD4.3 появляется журнал. 1, разряжены счетчики DD1 — DD3 в исходное состояние (рис. 192). В результате при выпуске элемента DD4.1 также появляется лог. 1 И следующий входной импульс отрицательной полярности устанавливает триггер DD4.2, DD4.3 в исходное состояние, сигнал сброса с входов R микросхемы DD1 — DD3 снимается и счетчик продолжает счет.
Спусковой механизм по элементам DD4.2 и DD4.3 обеспечивает сброс всех микросхем DD1 — DD3 при достижении счетчиком нужного состояния. При его отсутствии и больших порогах рассеяния микросхемы
DD1 — DD3 на входах R возможен случай, когда одна из микросхем DD1 — DD3 установлена в 0 и снимает сигнал сброса с других входов микросхемы раньше, чем сброс сигнал достигает порога их переключения. Однако такой случай маловероятен, и обычно можно обойтись без триггера, точнее без DD4.2 элемент.
Для получения коэффициента пересчета менее 10 для микросхемы К561И8 и менее 8 для К561И99 можно подключить выход дезинтектора с номером, соответствующим необходимому коэффициенту пересчета, напрямую с входом микросхемы R, например, как показано на рис. 193 (а) для коэффициента пересчета 6. Временный
Схема работы этого делителя показана на рис. 193 (6). Сигнал передачи может быть удален с выхода r только в том случае, если коэффициент пересчета равен 6 или больше для K561I8 и 5 или больше для K561I9.Для любого коэффициента сигнал передачи может быть удален с выхода декодера с номером, на единицу меньшего коэффициента пересчета.Индикацию состояния счетчиков счетчиков К176И8 и К561И8 удобно производить на газоразрядных индикаторах, согласовав их с помощью ключей на высоковольтных транзисторах NPN, например серии П307 — П309, КТ604, СТ605 или сборок К166Т1. (Рис.194).
Микросхемы K561Y10 и CR1561I10 (рис.195) содержат два отдельных четырехразрядных двоичных счетчика, каждый из которых имеет входы CP, CN, R. Установка триггеров счетчиков в начальное состояние происходит, когда журнал применяется к журналу r. 1. Логика работы входов CP и CN отличная от работы аналогичных микросхем К561И8 и микросхем К561И99. Триггеры микросхемы К561Ис и триггеры CR561I10 срабатывают по затуханию импульсов положительной полярности на входе КП с лог. 0 на входе CN (для K561I8 и K561I99 на входе CN должен быть лог.1) Возможны импульсы отрицательной полярности на входе CN, при этом Log 1 (для K561I8 и K561I9 для K561I8 и K561I9 — log 0) должен быть введен. Таким образом, входы CP и CN в микросхемах K561I10 и CR1561I10 объединены согласно элементной схеме, а в K561I8 и K561I9 — или.Временная диаграмма работы одного счетчика микросхемы свинца-Дена на рис. 196. При составлении микросхем в многозначный счет с последовательным переводом выходы 8 предыдущих счетчиков подключаются к входам далее CP, и бревно подается на входы CN.0 (рис.197). Если необходимо обеспечить параллельную передачу, следующие элементы and or or-no. На рис. 198 показана схема параллельного преобразования. Набор счетного импульса на входе счетчика CR DD2.2 через элемент DD1.2 допускается при состоянии 1111 счетчика DD2.1, когда он находится на выходе элемента DD3.1 log. 0. Аналогично прохождение счетного импульса на вход CP DD4.1 возможно только при состоянии 111 счетчиков DD2.1 и DD2.2 и т. Д. Назначение DD1.1 элемент такой же, как DD1.1 на рис. 189, и его можно исключить при тех же условиях. Максимальная частота входных импульсов для обоих вариантов счетчиков одинакова, но в счетчике с параллельной передачей переключение всех выходных сигналов происходит одновременно.
Из одной микросхемы счетчика можно построить делители частоты с коэффициентом деления от 2 до 16. Для примера на рис. 199 приведена схема коэффициента с коэффициентом пересчета 10 для получения коэффициентов пересчета -, 5, 6, 9, 12, можно использовать ту же схему, что и при соответствующем выборе выходов измерителя для подключения ко входам DD2.1 для получения коэффициентов пересчета 7, 11, 13, L4 Элемент DD2.1 должен иметь три входа, для коэффициента 15 — четыре входа.
Микросхема К561ИА11 — двоичный четырехразрядный реверсивный счетчик с возможностью параллельной записи информации (рис. 200). Микросхема имеет четыре информационных выхода 1, 2, 4.8, передающих передачу Р, и следующие входы: вход передачи ПИ, вход установки начального состояния R, вход подачи счетных импульсов с, вход счета счета U, входы для отправки информации с параллельной записью DL — D8, вводимой С.параллельно.Вход R имеет приоритет перед другими входами: если он на нем зарегистрирован. 1, на выходах 1, 2, 4, 8 будет лог.0 независимо от состояния других входов
. Если на входе r лог. 0 приоритет имеет вход S. При отправке журнала. 1 Асинхронная запись информации со входов D1 -D8 происходит в триггерах счетчика.Если на входах R, S, PI лог. 0, работа микросхем в счетном режиме разрешена. Если на входе u лог.1, при каждом спаде входного импульса отрицательной полярности, поступающего на вход C, состояние счетчика будет увеличиваться на единицу. С журналом. 0 На входе U Переключатели счетчика
В режиме вычитания — при каждом спаде импульса отрицательной полярности на входе состояние счетчика уменьшается на единицу. Если вы отправляете журнал на вход передачи PI. 1, режим подсчета запрещен.
На выходе передачи r log. 0, если на входе в PI log. 0 И все триггеры счетчиков находятся в состоянии 1 при увеличении счета или в состоянии 0 при неработающем счете.
Чтобы подключить микросхемы к последовательному счетчику передачи, вы должны объединить все входы с, выходы микросхем подключены к следующим входам следующих, и журнал должен представлять журнал на входе PI. 0 (рис.201). Выходные сигналы всех микросхем счетчика изменяются одновременно, однако максимальная частота счетчика меньше, чем у отдельной микросхемы из-за накопления задержек в схеме передачи. Чтобы обеспечить максимальную рабочую частоту многоразрядного счетчика, необходимо обеспечить параллельную передачу, для чего необходимо подать лог на входы ИП всех микросхем.Да, и сигналы на входы с микросхем подавать через дополнительные элементы или, как показано на рис. 202. В этом случае прохождение счетного импульса на входы с микросхемами будет разрешено только тогда, когда на выходах детали всех предыдущих чипов журнала. 0,
Причем время задержки этого разрешения после одновременного ответа микросхемы не зависит от количества разрядов счетчика.
Особенности построения микросхемы К561И1И11 требуют, чтобы входной сигнал направления на входе U возникал в паузе между счетными импульсами на входе С, то есть при лог.1 На этом входе или по снижению пульса.
Микросхема К176ИА12 предназначена для использования в электронных часах (рис. 203). В его состав входит кварц g G с внешним кварцевым резонатором на частоту 32768 Гц и два делителя частоты: ST2 на 32768 и ST60 на 60. При подключении к микросхеме кварцевого резонатора согласно рис. 203 (b) он обеспечивает частоты 32768, 1024, 128, 2, 1, 1/60 Гц. На выходах микросхемы Т1 — Т4 формируются импульсы частотой 128 Гц, колодец их равен 4, они сдвинуты на четверть периода.Эти импульсы предназначены для переключения выражения индикатора часов при динамической индикации. На счетчик-счетчик поступают импульсы с частотой 1/60 Гц, импульсы с частотой 1 Гц могут использоваться для подачи на второй счетчик и для обеспечения мигания точки отрыва импульсы с частотой 2 Hz можно использовать для установки показаний часов. Частота 1024 Гц предназначена для звукового сигнала тревоги и наблюдения за разрядами счетчиков с динамическим отображением, регулируется выходная частота 32768 Гц.Соотношения фаз колебаний разной частоты относительно момента снятия сигнала сброса показаны на рис. 204, временные масштабы различных диаграмм на этом шаблоне различны. При использовании
импульсов с выходов Т1 — Т4 для других целей следует обращать внимание на наличие коротких ложных импульсов на этих выходах.Особенность микросхемы в том, что первое спад на выходе минутных импульсов М появляется через 59 ° С. После снятия установочного сигнала 0 со входа R.Вызывает кнопку при запуске часов, кнопка генерирует сигнал установки 0, через одну секунду после шестого сигнала калибровки времени. Фронты и декали Сигналы на выходе y m синхронны с затуханиями импульсов отрицательной полярности на входе C.
Сопротивление резистора R1 может иметь такое же значение, как и для микросхемы К176ИА5. Конденсатор C2 служит для точной настройки частоты, C- для грубой. В большинстве случаев конденсатор С4 можно исключить.
Микросхема К176ИА13 предназначена для построения электронных часов с будильником.Он содержит счетчики минут и часов, регистр памяти будильника, цепочку сравнения и звуковой сигнал, схему динамической выдачи чисел для подачи на индикаторы. Обычно микросхема К176ИА13 используется совместно с К176ИА12. Стандартное подключение этих микросхем показано на рис. 205. Основными выходными сигналами схемы на рис. 205 являются импульсы Т1 — Т4 и числовые коды на выходах 1, 2, 4, 8. В моменты, когда на выходе Журнал T1. 1, на выходах 1,2,4,8 присутствует кодовое количество единиц минут при лог.1 На выходе Т2 — код количества десятков минут и т. Д. На выходе S — импульсы с частотой 1 Гц для зажигания точки отрыва. Выходные импульсы с служат для включения записи цифрового кода в регистр микросхемы C176ID2 или K176IID, обычно используются вместе с K176I12 и K176IA13, импульс на выходе на выход может использоваться для очистки индикаторов при корректировке показаний часов. Индексация индикаторов необходима, так как в момент коррекции происходит остановка динамической индикации и при отсутствии гашения горит только один разряд с крупнозернистой яркостью.На выходе ТГ — выход тревоги. Использование выходов S, K, HS не требуется. Журнал подачи. 0 Вход V-чипа переводит его выходы 1, 2, 4, 8 и c в состояние высокого нажатия.
При подаче питания на микросхемы счетчика часов минуты и нули автоматически записываются в регистр памяти сигналов тревоги. Для ввода в счетчик минут начального показания необходимо нажать
Кнопка SB1, показания счетчика начнут изменяться с частотой 2 Гц от 00 до 59 и затем снова 00, в момент перехода с 59 до 00 показания счетчика моточасов увеличатся на единицу.Показания счетчика часов также будут меняться с частотой 2 Гц от 00 до 23 и снова 00, если нажать кнопку SB2. Если вы нажмете кнопку SB3, на индикаторах появится время начала сигнала будильника. При одновременном нажатии кнопок SB1 и SB3 точки разряда за минуту включения будильника будут меняться от 00 до 59 и снова 00, но перехода в разряд не происходит. Если нажать кнопки SB2 и SB3, то разряд времени будильника изменится, при переходе из состояния 23 в 00 разрядятся показания.Вы можете нажать сразу три кнопки, в этом случае будут изменены показания как разряда минут, так и часов.Кнопка SB4 используется для запуска часов и коррекции хода во время работы. Если вы нажмете кнопку SB4 и отпустите ее через одну секунду после шестого сигнала калибровки времени, будут установлены правильные показания и точная фаза счетчика минут. Теперь вы можете выставить показания счетчика часов, нажав кнопку SB2, при этом ход счетчика не будет нарушен.Если показания счетчика находятся в пределах 00 … 39, показания счетчика часов при нажатии и отпускании кнопки SB4 не изменятся. Если показания минутного счетчика находятся в пределах 40 … 59, после отпускания кнопки SB4 показания счетчика часов увеличиваются на единицу. Таким образом, чтобы скорректировать ход часов, независимо от того, опоздали часы или торопились, достаточно нажать кнопку SB4 и отпустить ее через секунду после шестого сигнала калибровки времени.
Стандартная схема включения питания кнопок установки времени имеет тот недостаток, что при случайном нажатии кнопок SB1 или SB2 происходит сбой показаний часов.Если в размере рис. 205 добавить один диод и одну кнопку (рис. 206), то показания часов можно будет изменить, только нажав сразу две кнопки — кнопку SB5 («Установка
ка») и SB1. или кнопку SB2, что гораздо менее вероятно сделано случайно.Если показания часов и время включения сигнала тревоги не совпадают, на выходе HS микросхемы K176IA13 лог. 0. По совпадению появляются совпадения на выходе HS с положительной полярностью с частотой 128 Гц и длительностью 488 мкс (диета 16).Когда они передаются через ретранслятор эмиттера на любой эмиттер, сигнал напоминает звук обычного механического будильника. Сигнал транс-струнный, когда показания часов и будильника перестают совпадать.
Схема согласования микросхемы CC176i12 и K176IA1313 с индикаторами зависит от их типа. Например, на рис. 207 В зависимости от схемы подключения полупроводниковых семипозиционных индикаторов с общим анодом. Как катодный (VT12 — VT18), так и анодный (VT6, VT7, VT9, VT10) ключи выполнены по схемам эмиттерных повторителей.Резисторами R4 — R10 определяется импульсный ток через отрезки индикаторов.
Указанное на рис. 207 Удельное сопротивление резисторов R4 -R10 обеспечивает импульсный ток через сегмент около 36 мА, что соответствует среднему току 9м. При таком токе индикаторы AL305A, ALS321B, ALS324B и другие имеют достаточно яркий ориентир. Максимальный коллекторный ток транзисторов VT12 — VT18 соответствует току одного сегмента 36 мА и поэтому здесь можно использовать практически любые маломощные транзисторы П-Н-П с разрядностью коллектора 36 мА и более.
Импульсные токи транзисторов анодных ключей могут достигать 7 x 36-252 мА, поэтому транзисторы, обеспечивающие указанный ток, могут использоваться в качестве анодных ключей, с коэффициентом передачи базового тока h31E не менее 120 (серия CT317, КТ503, КТ815).
Если нельзя подобрать транзисторы с таким коэффициентом, можно использовать составные транзисторы (CT315 + KT503 или KT315 + KT502). Транзистор VT8 представляет собой любую маломощную структуру N-P-N.Транзисторы VT5 и VT11 — Эмиттерные повторители для подключения излучателя звука будильника на 1, в котором могут использоваться любые телефоны, в том числе малогабаритные от слуховых аппаратов, любые динамические головки, включаемые через выходной трансформатор от любого радиоприемника.Подбором емкости конденсатора С1 можно добиться по необходимой громкости звукового сигнала, также можно установить переменный резистор 200 … 680 Ом, включая его потенциометр между С1 и 1. Переключатель SA6 служит для отключения сигнал тревоги.
Если используются общие катодные индикаторы, то эмиттерные повторители, подключенные к выходам микросхемы DD3, должны быть выполнены на транзисторах N-P-N (серия CT315 и другие серии), а вход S DD3 подключен к общему проводу.Для подачи импульсов на катоды. Индикаторы должны собирать ключи на транзисторах N-P-N по схеме с общим эмиттером. Их базы следует подключить к выходам Т1 — Т4 микросхемы DD1 через резисторы 3.3 ком. Требования к транзисторам такие же, как и к транзисторам анодных ключей в случае индикаторов с общим анодом.
Индикация возможна и с люминесцентными индикаторами. В этом случае необходима подача импульсов Т1 — Т4 на сеточные индикаторы и подключение интегрированных объектов индикаторов через одноименную микросхему К176ИМ2 или К176ИД на выходы 1, 2, 4, 8 микросхемы. K176IA13.
Схема подачи импульсов на индикаторных сетках представлена на рис. 208. Сетки С1, С2, С4, С5 — соответственно сетка знакомств единиц и десятков минут, единиц и десятков часов, С- сетка разделения точка. Индикаторы. Аноды должны быть подключены к выходам микросхемы K176IID, подключенной к DD2 согласно включению DD3 на рис. 207 с помощью клавиш, подобных нажатию клавиш. 178 (б), 179,180, ввод микросхемы К176ИМ2 должен быть подан. один.
Возможно использование микросхемы К176-без ключа, ее вход должен быть подключен к общему проводу.В любом случае аноды и сетчатые индикаторы должны быть через резисторы 22 … 100 кОм подключены к источнику отрицательного напряжения, которое находится по абсолютной величине от 5 … 10 до более отрицательного напряжения, которое суммировалось до катоды индикаторов. На схеме. 208 Это резисторы R8 — R12 и напряжение -27 В.
Перенос импульсов Т1 — Т4 на сетку индикаторов удобно осуществлять с помощью микросхемы К161КН2, подавая на нее напряжение питания согласно рис.180.В качестве индикаторов могут использоваться любые одинарные вакуумные люминесцентные индикаторы, а также плоские четырехместные индикаторы с разделительными точками ИВЛ1 — 7/5 и ИВЛ2 — 7/5, специально разработанные для часов. В качестве схемы DD4 рис. 208 можно использовать любые инвертирующие логические элементы с комбинированными входами.
На рис. 209 приведена схема согласования с газоразрядными индикаторами. Анодные ключи могут быть выполнены на транзисторах серии КТ604 или СТ605, а также на транзисторах сборки К166Т1.
Неоновая лампа HG5 служит для обозначения точки разделения. Индикаторные катодные затравки следует объединить и подключить к выходам декодера DD7. Для упрощения схемы можно исключить инвертор DD4, обеспечивающий срабатывание индикаторов в момент нажатия кнопки коррекции.
Возможность перевода выходов микросхемы K176IA13 в высокоимпедансное состояние позволяет построить часы с двумя показаниями (например, MSK и GMT) и двумя будильниками, один из которых может использоваться для включения любого устройства, другой для выключения (рис.210).
Входы основного DD2 и дополнительной микросхемы DD2 K176IA13 соединены между собой и с другими элементами согласно рис. 205 (можно опираться на рис. 206), за исключением входов P и V. На верхней схеме положение переключателя SA1
Установки с SB1 — кнопки SB3 могут входить на вход микросхемы DD2, в внизу — на DD2. Поток сигналов на микросхеме DD3 управляется секцией переключателя SA1.2. В верхнем положении горох за доплату sa1 log.1 поступает на вход V микросхемы DD2 и сигналы с выходов DD2. В нижнем положении переключатель лаг. 1 На входе V микросхемы DD2 разрешена передача сигналов с ее выходов.В результате в верхнем положении переключателя SA1 вы можете управлять первыми часами и будильником и отображать их состояние в нижней секунде.
Срабатывание первого будильника включает триггер DD4.1, DD4.2, появляется лог на выходе DD4.2. 1, который можно использовать для включения любого устройства, срабатывание второй сигнализации закрывает это устройство.Кнопки SB5 и SB6 также можно использовать для его включения и выключения.
При использовании двух микросхем K176IA13 Сигнал сброса на вход R микросхемы DD1 должен сниматься непосредственно с кнопки SB4. В этом случае происходит коррекция показаний, как показано на рис. 205 Подключение, но блокировка SB4 «Корр.» Кнопка
Нажатие кнопки SB3 «Бутон.» (Рис. 205), существующей в стандартной версии, не встречается. При одновременном нажатии кнопок SB3 и SB4 в часах с двумя микросхемами K176IA13 показания не работают, но не по ходу часов.Правильные показания восстанавливаются, если повторно нажать кнопку SB4 при отпускании SB3.Микросхема К561ИА14 — двоичная и двухзначная десятичная четырехзначная оценка-цыпленок (рис. 211). Его отличие от микросхемы К561И1И11 заключается в замене записи R на вход in — вход переключения модуля счета. С журналом. 1 На входе в микросхему К561II14 бинарный счет выдает, как и К561И11, с логом. 0 На входе — двоично-десятичный. Назначение остальных входов, режимы работы и правила включения у этой микросхемы такие же, как у К561И11.
Микросхема Ca561Is15 — делитель частоты с переключаемым коэффициентом деления (рис. 212). Микросхема имеет четыре входа управления KL, K2, K-, L, для подачи тактовых импульсов, шестнадцать входов для установки коэффициента деления 1-8000 и один выход.
Микросхема позволяет иметь несколько вариантов установки коэффициента деления, его диапазон колеблется от 3 до 21327. Друг будет считаться самым простым и удобным вариантом, для которого, однако, максимально возможный коэффициент деления составляет 16659.Для этого варианта необходимо постоянно применять Журнал. 0.Вход К2 используется для установки начального состояния счетчика, которое возникает в трех периодах входных импульсов, когда журнал применяется к входу. 0. После подачи бревна. 1 На входе К2 начинается работа счетчика в режиме частотного деления. Коэффициент частотного деления при отправке журнала. 0 На входах L и K1 10 000 и не зависит от сигналов, подаваемых на входы 1-8000. Если входы L и K1 подают разные входные сигналы (лог.0 и log 1 или log. 1 и log 0) коэффициент деления входной частоты импульсов определяется двоично-десятичным кодом, подаваемым на входы 1-8000. Например, на рис. 213 Временная диаграмма микросхемы показана в режиме деления на 5, для обеспечения которого необходимо подать журнал на входы 1 и 4. 1, входы 2, 8-8000 — лог. 0 (K1 не равно l).
Длительность выходных импульсов положительной полярности равна периоду входного импульса, фронты и спады выходных импульсов совпадают с затуханиями входных импульсов отрицательной полярности.Как видно из временной диаграммы, первый импульс на выходе микросхемы появляется на затухании входного импульса с номером, приходящимся на единицу большого коэффициента деления.
При отправке журнала. 1 На входах L и K1 выполняется единый учет. При подаче на вход К2 лог. 0 На выходе микросхемы появляется лог. 0. Длительность начального установочного импульса на входе К2 должна быть, как и в режиме частотного разделения, не менее трех периодов входных импульсов.После окончания на входе начальный установочный импульс начнет учет, который будет происходить по спаду входных импульсов отрицательной полярности. После окончания импульса с номером, единица большим кодом, установленным на входах 1-8000, лог. 0 Вывод изменится в журнале. 1, после чего не будет изменен (рис. 213, К1 — Л — 1). Для следующего запуска необходимо ввести начальный установочный импульс для входа в K2.
Данный режим микросхемы аналогичен работе ожидающего мультивибратора с установленной длительностью цифрового импульса, следует только помнить, что длительность входного импульса включает в себя длительность исходного установочного импульса и, более того, другой период входных импульсов. .
Если после окончания формирования выходного сигнала в режиме единого счета ввести К1 для подачи журнала. 0 микросхема переключится в режим деления входной частоты, а фаза выходного импульса будет определяться начальным установочным импульсом, поданным ранее в режиме единого счета. Как упоминалось выше, микросхема может обеспечить фиксированный коэффициент деления частоты, равный 10 000, если входные данные журнала подаются в журнал. 0. Однако после начального установочного импульса, подаваемого на вход К2, после входа появляется первый выходной импульс с импульсом с номером на единицу большого кода, установленного на входах 1-8000.Все последующие выходные импульсы появятся через 10 000 периодов входных импульсов после начала предыдущего.
На входах 1-8 допустимые комбинации входных сигналов должны соответствовать двоичному эквиваленту десятичных чисел от 0 до 9. На входах 10-8000 разрешены произвольные комбинации, то есть можно подавать числа от 0 до 15 каждую декаду. В результате максимально возможный коэффициент деления будет:
К — 15000 + 1500 + 150 + 9 = 16659.
Микросхема может быть использована в синтезаторах частот, электроинструментах, программируемых реле времени, для формирования точных временных интервалов в различных устройствах.
Микросхема К561Ис16 представляет собой четырнадцатиразрядный двоичный счетчик с последовательной передачей (рис. 214). Микросхемы имеют два входа-входа установки начального состояния R и вход подачи тактовых импульсов S. Установка триггеров счетчика в 0 производится при подаче логгера на вход R.1, счет — по спадам импульсов положительной полярности, подаваемых на вход S.Счетчик не имеет выходов всех разрядов — нет выходов разрядов 21 и 22, поэтому при необходимости иметь сигналы с Для всех разрядов двоичного счетчика следует использовать другой счетчик, работающий синхронно и имеющий выходы 1, 2, 4, 8, например половину микросхемы K561Is (рис. 215).
Коэффициент деления одной микросхемы К561У16 равен 214 = 16384, при необходимости больший коэффициент отбора можно освободить 213 микросхем для совмещения с входом другой такой же микросхемы или с входом КП любой другой микросхемы — прилавок.10 предыдущего, можно уменьшить разряд счетчика для получения недостающих выходов двух цифр второй микросхемы (рис. 216). Подключив вход микросхемы K561Y1I к микросхеме K561I10, можно не только получить недостающие выходы, но и увеличить разряд метра на единицу (рис. 217) и обеспечить коэффициент деления 215 = 32768.K561Y1I16 Микросхема удобно использовать в делителях частоты с перестроенным коэффициентом деления по схеме, подобной рис.3, следует использовать схему. 215 или 59, с коэффициентом более 16384 — схема рис. 216.
Для перевода числа в двоичную форму, его нужно разделить на 2, остаток (0 или 1) записать. Полученный результат снова делится на 2, остаток записывается и так до тех пор, пока после деления не останется ноль. Первый остаток — младший разряд двоичной формы числа, последний — старший.
Микросхема К176ИА17 — Календарь. Он содержит счетчики дней недели, числа месяца и месяцев.Счетчик чисел считает от 1 до 29, 30 или 31 в зависимости от месяца. День недели — от 1 до 7, счет месяцев — от 1 до 12. Схема микросхемы К176ИА на микросхему К176ИА 13 часов представлена на рис. 219. На выводах 1-8 микросхемы DD2 расположены поочередно. цифры и коды месяцев аналогичны часам и минутам на выходах
Микросхема К176ИА13. Подключение индикаторов к указанной молодежной микросхеме К176ИА17 выполнено аналогично им для подключения к выходам микросхемы К176ИА с использованием записывающих импульсов с выхода микросхемы К176ИА13.На выходах A, B, с постоянно присутствующим кодом 1-2-4 порядкового номера дня недели. Его можно применить к микросхеме К176ИМ2 или К176ИД, а затем и на любом семисегментном индикаторе, в результате чего на нем будет отображаться номер недели. Однако более интересным является возможность вывода двухбуквенного обозначения дня недели на цифровые индикаторы ИВ-4 или ИВ-17, для чего необходимо сделать специальный преобразователь кодов.
Установка числа, месяца и дня недели производится аналогично установке показаний в микросхеме К176II13. При нажатии кнопки SB1 устанавливается номер, кнопка SB2 — месяц, при свернутом SB3 и SB1 — день недели. Для уменьшения общего
Номера кнопок в часах с календарем можно использовать схемы SB1 -SB3, SB5. 206 Для настройки индикаторов календаря переключение их общей точки на тумблер с входа микросхемы K176IA13 на вход P микросхемы K176IA17.Для каждой из этих микросхем схема R1C1 должна быть аналогична схеме. 210.Журнал подачи. 0 Вход микросхемы переводит его выходы 1-8 в состояние с высоким импедансом. Это свойство микросхемы позволяет относительно легко организовать поочередную выдачу часов и календаря на один четырехзначный индикатор (кроме дня недели). Схема
Подключение микросхемы K176ID2 (ID-3) к микросхемам IE13 и IE17 для обеспечения указанного режима показано на рис. 220, составные схемы микроскопа K176IA13, Ye17 и Ye12 друг с другом не показаны.В верхней схеме положения переключателя SA1 («часы») выходы 1-8 микросхемы DD3 находятся в состоянии high imaging, выходные сигналы микросхемы DD2 через резисторы R4 — R7 поступают на входы DD4. входы микросхемы, состояние микросхемы DD2 — отображаются часы и минуты. В нижнем положении переключателя SA1 («календарь») выходы микросхемы DD3 активируются, и теперь микросхема DD3 определяет входные сигналы микросхемы DD4. Переведите выходы микросхемы DD2 в состояние высокого напряжения, как это сделано в схеме
Рис.210, невозможно, так как он перейдет в высокоимпедансное состояние и будет выводиться с микросхемы DD2, а аналогичный вывод микросхемы DD3 не имеет. На схеме. 220 Реализовано выше с помощью одного набора кнопок для установки показаний часов и календаря. Импульсы от кнопок SB1 — SB3 поступают на вход P микросхемы DD2 или DD3 в зависимости от положения того же переключателя SA1.Микросхема К176ИА18 (рис. 221) во многом напоминает К176И12. Его основные отличия — исполнение выходов Т1 — Т4 с открытым ложем, что позволяет подключать к этой микросхеме сетки вакуумных люминесцентных индикаторов без согласования ключей.
Для обеспечения надежной фиксации индикаторов на своих сетках скважность импульсов Т1 — Т4 в микросхеме К176ИА18 составляет чуть больше четырех и составляет 32/7. При отправке журнала. 1 на входе R микросхемы на выходах Т1 — Т4 лог. 0, поэтому подача специального сигнала пожаротушения на ввод микросхемы К176ИИД2 и К176ИД3 не требуется.
Вакуумные люминесцентные индикаторы зеленого свечения в темноте кажутся значительно ярче, чем на свету, поэтому желательно иметь возможность изменять яркость индикатора.Микросхема К176ИА18 имеет вход Q, питающий бревно. 1 к этому входу можно в 3,5 раза увеличить количество импульсов на выходах Т1 — Т4 и в
За то же время уменьшить яркость свечения индикаторов. Сигнал на вход Q может подаваться либо с переключателя яркости, либо с фоторезистора, второй выход которого подключен к плюсу питания. В этом случае Q в этом случае следует подключить к общему проводу через резистор 100 КОМ … 1 МОм, который необходимо выбрать для получения желаемого порога внешнего освещения, при котором будет происходить автоматическое переключение яркости.Следует отметить, что когда лог. 1 На входе Q (Малая яркость) Не работает установка показаний часов.
Микросхема К176ИА18 имеет специальный генератор звукового сигнала. При подаче импульса положительной полярности на вход ГС на выходе ГС возникают импульсы отрицательной полярности с частотой 2048 Гц и длительностью 2. Длительность пачек — 0,5 с, период следования 1 с. Выход HS выполнен с открытым прикладом и позволяет подключать эмиттеры с сопротивлением 50 Ом и выше между этим выходом и плюсом питания без репитера эмиттера.Сигнал присутствует на выходе HS до конца очередного минутного импульса на выходе микросхемы.
Следует отметить, что допустимый выходной ток микросхемы К176ИА18 по выходам Т1 — Т4 составляет 12 мА, что значительно превышает ток микросхемы К176ИА12, следовательно, требования к усилению транзисторов в ключах при применении К176ИА18 микросхемы и полупроводниковые индикаторы (рис. 207) значительно менее жесткие h31E> 20. Базовое сопротивление
Резисторы в катодных ключах можно уменьшить до 510 Ом при h31E> 20 или до 1Q0M при h31E> 40.
Микросхемы К176ИА12, К176ИА13, К176ИА17, К176ИБ18 Регулировка напряжения питания такая же, как у микросхем серии К561 — от 3 до 15 В.
Микросхема К561ИА19 представляет собой пятиразрядный регистр сдвига с возможностью параллельной записи информации, предназначен для построения счетчиков с программируемым модулем учета (рис. 222). Микросхема имеет пять информационных входов для параллельной записи D1 -D5, входную информацию для последовательной записи DO, вход параллельного входа S, вход сброса R, вход для подачи тактовых импульсов и пять обратных выходов 1-5.Вход R является преобладающим — при отправке журнала. 1 Все триггеры микросхемы выставлены на 0, лог появляется на всех выходах. 1 Независимо от сигналов на других входах. При подаче на вход R log. 0, в журнале регистрации. 1 Информация записывается со входов D1 — D5 в триггеры микросхемы, на выходах 1-5 появляется в обратном виде.
При подаче на входы R и S лог. 0 Возможен сдвиг информации в триггерах микросхемы, который будет происходить при затуханиях импульсов отрицательной полярности, поступающих на вход C.В первом триггере информация будет записана со входа D0.
Если соединить вход DO с одним из выходов 1-5, можно получить счетчик с коэффициентом пересчета 2, 4, 6, 8, 10. Для примера на рис. 223 Временная диаграмма микросхемы отображается в режиме деления на 6, который организован в случае соединения входа D0 с выходом 3. Если вам нужно получить нечетный коэффициент пересчета 3.5.7 или 9, вы должны использовать двусторонний элемент и , входы которого подключены соответственно к выходам 1 и 2, 2 и 3, 3 и 4.4 и 5, выход на вход DO. Например, на рис. 224 показана схема делителя частоты для 5, на рис. 225 — временная диаграмма его работы.
Следует иметь в виду, что использование микросхемы К561Ис19 невозможно в качестве сдвигового регистра, так как он содержит корректирующие цепочки, в результате чего совмещение состояний триггеров, не являющихся рабочими для режима счета исправляются автоматически. Наличие схем коррекции позволяетАналогично использованию микросхем K561Y8 и K561I9 Не подавайте начальный установочный импульс на счетчик, если фаза выходных импульсов не важна.12 = 4096. Имеет два входа — R (для установки нулевого состояния) и с (для подачи тактовых импульсов). С журналом. 1 На входе R счетчик установлен в нулевом состоянии, а с логом. 0 — считает спад положительной полярности на вход от импульсов. Микросхему можно использовать для разделения частоты на коэффициенты, которые являются степенью числа 2. Чтобы построить делители с другим коэффициентом деления, вы можете использовать схему, включающую микросхему K561Y1I (рис. 218).
Микросхема КР1561И21 (рис.227) — синхронный двоичный счетчик с возможностью параллельной записи информации о спаде тактового импульса. Принцип работы микросхемы аналогичен К555У10 (рис. 38).
Понятен принцип работы К176ИА4. В этой статье я хочу рассказать о принципе работы с K176IA4 — незаменимым драйвером семисегментных индикаторов. Предлагаю разобрать его работу на примере этой схемы: не бойтесь — хоть и выглядит массивно, несмотря на это очень просто, используется всего 29 электронных компонентов по принципу работы К176И4: К176И4 — по сути очень простая в понимании микросхема.Это десятичный счетчик с декодером семисегментной индикации. Имеет 3 входа и 9 сигнальных выходов. Номинальное напряжение питания — от 8,55 до 9,45 В. Максимальный ток на один выход — 4 входа это: тактовая линия (4 ножки микросхемы) — она доходит до нее, что заставляет микросхему переключать свои состояния, то есть рассмотреть выбор общего анода / катода (6 ножек) — подключив эту линию к минусу, мы можем погонять индикатор с общим катодом, к плюсу — с общим сбросом анода (5 ножек) — при подаче лога.1 сбрасывает счетчик на ноль при отправке журнала. 0 — позволяет микросхеме переключать выходы состояний: 7 выходов на семиступенчатый индикатор (1, 8-13 ножки) тактирующий сигнал, разделенный на 4 (3 ножки) — необходим для тактовых цепей, мы не используем тактирующий сигнал, деленный на 10 (2 ноги) — позволяет Комбинировать несколько К176ИА4, расширяя диапазон разрядов (можно добавить десятки, сотни и т. Д.) Принцип расчета работает таким образом, что при переключении сигнала на тактовую линию с логом . 0 в журнале.1 Значение тока увеличивается по принципу действия этой схемы: Для упрощения восприятия работы этой схемы можно сделать такую последовательность: NE555 выдает прямоугольный импульс K176IA4 под действием импульса увеличивает свое состояние к одному. Его текущее состояние передается на сборку транзисторов ULN2004 для усиления усиленного сигнала. Индикатор отображает текущий статус. Эта схема переключает состояние YE4 один раз в секунду (этот промежуток времени формируется цепочкой RC, состоящей из R1, R2 и C2) NE555 можно спокойно заменить на KR1006V1 C3, вы можете выбрать в диапазоне от 10 до 100 NF, усилитель необходим, так как максимальный ток на выходе IE4 — 4 мА, а номинальный ток большинства светодиодов — 20 мА. Семиступенчатые индикаторы подойдут для любых с общим анодом и номинальным напряжением от 1.От 8 до 2,5В, при токе от 10 до 30мА подключаем 6 ножку микросхемы к минусу питания, но используем индикатор с общим анодом, это связано с тем, что ULN2004 не только усиливает, но и инвертирует микросхему сигнал сбрасывает свое состояние при подаче питания (по схеме C4 и R4) или при нажатии кнопки (S1 и R3). Сброс при подаче питания. Поскольку в противном случае микросхема не будет нормально работать, резистор до кнопки сброса необходим для безопасной работы кнопки — почти все кнопки часов рассчитаны на ток не более 50мА, поэтому резистор надо выбирать из 9В / 50MA = 180 и до 1ку Автор: Arssev1 Взято с http: // cxem.нетто 20 шт. NE555 NE555P NE555N 555 DIP-8. Цена FOB для Справки: US $ 0.99 / Партия
На последнем занятии мы познакомились с микросхемой К561И8, содержащей десятичный счетчик и десятичный десятичный разряд, а также с микросхемой К176ИИД, содержащей декодер, рассчитанный на работу с семью индикаторами. Существуют микросхемы К176 и К176ИА4, содержащие счетчик и декодер, рассчитанные на работу с индикатором семерки.
Чипы имеют одинаковые соединения и корпуса (показаны на рис. 1A и 1B на примере микросхемы K176IA), разница в том, что K176 подчиняется 6, а K176I4 — 10.Микросхемы предназначены для электронных часов, поэтому K176 сообщает, например, 6, если вам нужно считать десятки минут или секунд. Кроме того, обе микросхемы имеют дополнительный вывод (вывод 3). В микросхеме К176II4 единица появляется на этом выходе в момент, когда ее счетчик переходит в состояние «4». А в микросхеме k176 на этом выходе появляется один в тот момент, когда счетчик занимает до 2. Таким образом, наличие этих выводов дает возможность построить счетчик часов, считая до 24.
Рассмотрим микросхему K176IA4 (рис. 1A и 1B). На вход «С» (выход 4) подаются импульсы, которые микросхема должна считать и отобразить их количество в семикратной форме на цифровом индикаторе. Вход «R» (выход 5) служит для принудительной установки счетчика микросхемы в ноль. Когда к нему применяется логическая единица, счетчик переходит в нулевое состояние, а на индикаторе, подключенном к рендерингу оборудования чипа, отображается цифра «0», выраженная в семисегментной форме (см. Урок No.9). Счетчик микросхемы имеет передачу «П» (вывод 2). 10 на этом выходе микросхема считает логической единицей. Как только микросхема достигает 10 (на ее входе десятый импульс), она автоматически возвращается в нулевое состояние, и в этот момент (между спадом 9-го импульса и фронтом 10-го) на выходе « р »на выходе формируется отрицательный импульс (нулевой дифференциал). Наличие этого выхода «P» позволяет использовать микросхему в качестве делителя частоты на 10, поскольку частота импульсов на этом выходе будет в 10 раз ниже, чем частота импульсов, поступающих на вход «C» (каждые 10 импульсов при вход «С» — на выходе «П» — один импульс).Но основная цель этого выхода («П») — организация мерного счетчика.
Другой вход — «s» (выход 6), он нужен для выбора типа индикатора, с которым будет работать микросхема. Если это светодиодный индикатор с общим катодом (см. Урок № 9), то для работы с ним на этот вход нужно подать логический ноль. Если индикатор с общим анодом — представить единицу.
Выходы «A-G» используются для управления сегментами светодиодного индикатора, они подключены к соответствующим входам семиконтактного индикатора.
Микросхема мез К176 работает так же, как и К176ИА4, но учитывает только 6, а на ее выходе 3 появляется одна, когда счетчик принимает до 2. В остальном микросхема не отличается от К176.
Для исследования микросхемы К176ИА4 собирают схему, показанную на рисунке 2. На микросхеме Д1 (К561Л5 или К176Л5) встроен формирователь импульсов. После каждого нажатия и отпускания кнопки S1 на ее выходе (на выходе 3 d1.1) формируется один импульс. Эти импульсы поступают на вход «с» микросхемы D2 — К176И4.Кнопка S2 служит для подачи единичного логического уровня на вход «R» D2 для перевода, таким образом, счетчика микросхемы в нулевое положение.
Светодиодный индикатор h2 подключен к выходам микросхемы A-G. В этом случае используется индикатор с общим анодом, поэтому для зажигания его сегментов на соответствующих выходах D2 должны быть нулями. Для перевода микросхемы D2 в режим работы с такими индикаторами на ее вход S (выход 6) подается питание.
С помощью вольтметра Р1 (тестер, мультиметр включен в режим измерения напряжения) можно наблюдать изменение логических уровней при переходе передачи (вывод 2) и на выходе «4» (вывод 3). .
Установите микросхему D2 в нулевое состояние (нажмите и отпустите S2). Индикатор h2 покажет цифру «О». Затем, нажав кнопку S1, проследите за работой счетчика от «0 до« 9 », и при следующем нажатии снова перейдет к« 0 ». Затем установите диаграмму прибора на выход 3 d2 и нажмите S1. Сначала , при этом на счету идет от нуля до трех до трех. Этот выход будет нулевым, но с появлением числа «4» — на этом выходе будет блок (устройство R1 покажет напряжение, близкое к напряжению питания).
Попробуйте соединить выводы 3 и 5 микросхемы D2 при помощи отрезка монтажного провода (на схеме изображен штрих-код). Теперь счетчик, достигший нуля, будет принимать только «4». То есть показатели индикатора будут такими — «0», «1», «2», «3» и еще раз «0» и далее по кругу. Вывод 3 позволяет ограничить счет фишки до четырех.
Установите датчик прибора P1 на выход 2 d2. Все время прибор будет показывать единицу, но после 9-го импульса в момент получения 10-го импульса и перехода в ноль уровень упадет до нуля, а затем, после десятого, снова станет одинокий.Используя этот выход (выход P), вы можете организовать многоразрядный счетчик.
На рисунке 3 представлена схема двухразрядного счетчика, построенного на двух микросхемах К176ИА4. Импульсы на вход этого счетчика поступают от выхода мультивибратора на элементах D1.1 и D1.2 микросхемы К561Л5 (или К176Л5).
Счетчик на D2 считает единицы импульсов, и после каждых десяти импульсов, поступивших на его вход «с», на его выходе «P» появляется один импульс. Второй счетчик — D3 считает эти импульсы (поступающие с выхода «p» счетчика D2), а его индикатор показывает десятки импульсов, поступивших на вход D2 с выхода мультивибратора.
Таким образом, этот двузначный счетчик считает от «00» до «99» и с приходом сотого импульса обнуляется.
Если нам нужен этот двузначный счетчик на and39 «(передается в ноль при получении 40-го импульса), нужно вывести 3- d3 с помощью отрезка монтажного провода для соединения с соединенными вместе выводами 5 обоих счетчиков. Теперь, когда закончится третья палатка входных импульсов, блок с выхода 3 -D3 перейдет на «R» входы обоих счетчиков и обнулит их.
Для изучения микросхемы мехов К176 соберем схему, показанную на рисунке 4.
Схема такая же, как на рисунке 2. Разница в том, что микросхема будет считать от «примерно» до «5», а при получении 6-го импульса до нулевого состояния. На выходе 3 появится единица при входе на второй импульсный вход. Импульс передачи на выходе 2 появится с приходом 6-го входного импульса. Пока что считает до 5 на выходе 2 — единица, с приходом 6-го импульса в момент перехода в ноль — логический ноль.
Используя две микросхемы К176 и К176ИА4, можно построить счетчик по аналогии с электронными часами для отсчета секунд или минут, то есть счетчик считает до 60. На рисунке 5 представлена схема такого счетчика.
Схема такая же, как на рисунке 3, но разница в том, что в качестве микросхемы D3 вместе с K176I4 используется K176. И эта микросхема считает до 6, тогда количество десятков будет 6. Счетчик будет считать от «00» до «59», а с приходом 60-го импульса уйти в ноль.Если сопротивление резистора R1 выбрано таким, чтобы импульсы на выходе D1.2 следовали с периодом в одну секунду, можно получить секундомер, который работает до одной минуты.
С помощью этих микросхем легко построить электронные часы.
Это будет наше следующее занятие.
Журнал Radio Constructor 2000
Дополнительно
Кейс: ДИП-14
Микросхема К176ИА4 Представляет собой счетчик модуля 10 с декодером для вывода информации на индикатор семерки.Микросхема К176ИА4 была разработана специально для работы в схемах электронных часов.
Учет происходит по затуханию импульсов положительной полярности на тактовом входе С. Подача лога. «1» на входе R переводит триггеры счетчика в нулевое состояние. Вход S контролирует «полярность» сигналов на выходах сегментов — это позволяет использовать индексы как с общим анодом, так и с общим катодом.
На выходе 2 выдается последовательность импульсов с частотой F / 10, на выходе 3 — f / 4.
Аналог: CD4026B.
Обозначения K176IA44:
Назначение выводов К176ИА4:
Несмотря на то, что серия K176 относится к устаревшей CMOS-серии, некоторые микросхемы этой серии, в частности K176IA4, не имеют аналогов в более современных сериях K561 / CR561 и поэтому все еще востребовано в отдельных приложениях.
Основные параметры K176II4:
Подключение ЖК-индикатора к K176II4:
Подключение люминесцентного индикатора к К176ИА44:
Схемы подключения светодиодных индикаторов к К176И4:
Действие цифрового частотомера основано на измерении количества входных импульсов в течение примерного временного интервала в 1 секунду.
Исследуемый сигнал поступает на вход генератора импульсов, который собран на транзисторе VT1 и элементе DD3.1, который производит электрические колебания прямоугольной формы, соответствующие частоте входного сигнала.
Технические характеристики
- Время измерения, Кл — 1
- Максимальная измеряемая частота, Гц — 9999
- Амплитуда входного сигнала, в — 0,05 … 15
- Электропитание, в — 9.
Принципиальная схема
Эти импульсы поступают на электронный ключ DD3.2. На другой ключевой вход (выход 5 DD3.2) от управляющего устройства принимаются импульсы с частотой дискретизации, удерживая клавишу открытой в течение 1 секунды.
В результате на ключевом выходе (выход 4 элемента DD3.2) формируются пучки импульсов, которые поступают на вход счетчика DD4 (выход 4).
Рис. 1. Принципиальная схема цифрового частотомера на микросхемах.
Примерный генератор частоты (рис. 1) собран на микросхеме DD1 и кварцевом резонаторе zQ1.Импульсы от него поступают на управляющее устройство, представляющее собой D-триггер DD2. Триггер делит тактовую частоту на две части.
Фронт входного импульса переводит триггер в единичное состояние. Происходит кратковременный сброс счетчиков DD4 … DD7. Транзистор VT2 принимает сигнал низкого уровня и закрывает его, поэтому индикаторы HL1 … HL4 зацепляются. Допускается работа клавиши DD3.2, и импульсы поступают на вход счетчика.
Другой импульс частоты дискретизации переводит триггер DD2 в нулевое состояние.Клавиша DD3.2 закрывается. Сигнал высокого уровня с выхода 2 микросхемы DD2 открывает транзистор VT2 и включает индикаторы HL1 … HL4, которые отображаются для результата измерения в течение 1 секунды.
Детали
В схеме используется кварц ZQ1 на частоту 32768 Гц. Микросхему К176ТМ2 и К176Л7 можно заменить на К561ТМ2 и К561Л7 соответственно. Вместо К176ИА12 можно применить К176ИА5, с соответствующей корректировкой схемы.
Следующая схема счетчика является простейшим примером использования микросхем К176ИА4, представляющих собой десятичные счетчики с декодером.
На микросхеме создан импульсный генератор для переключения счетчиков. Резистор R1 и конденсатор С1 (в основном резистор) Частота импульсов задается. С такими элементами, как на схеме, частота была получена 1,2 с.
К176ИА4 — Счетчик импульсов с выводом состояния счетчика на семиступенчатый индикатор. Считает импульсы, поступившие на вход из (4-х ног). По затуханию этих импульсов счетчик переключается. На выходе «J» (3 микросхемы) частота в 4 раза меньше тактовой частоты, а на выходе «P» (2 микросхемы) частота в 10 раз меньше тактовой частоты на нем, спад логическая единица возникает, когда состояние счетчика переходит с «9» в «0».Он используется для подключения следующего счетчика высшей категории. Вход R служит для сброса счетчиков, происходит при появлении на нем логической единицы. Следует отметить, что если эта запись висит в воздухе, то связи ни с чем нет, микросхема часто там блок воспринимает, а счет не производит. Во избежание этого необходимо подтянуть его к земле, подключив на общий минус через резистор 100 — 300 Ом, либо напрямую, если не планируется использовать функцию нуля.Вход S предназначен для переключения режимов микросхемы с разными показателями. Если к этому выводу подключено + питание, микросхема переходит в режим работы с индикатором с общим анодом, если C — питание находится в режиме индикатора с общим катодом. Выходы 1, 8 — 13 служат для подключения индикатора.
IC1 учитывает импульсы генератора, поступившие на его вход 4, при переключении его с 9 на 0 на выходе 2 логическая единица уменьшается, а IC2 переключается на 1 значение вверх.
Клавиша S1 управляет питанием, S2 сбрасывает счетчики (вместо этого я использовал зародыш и магнит).
Для индикатора требуется семисегментный двусторонний (или два семисегментных индикатора). Если индикатор с общим катодом (минус), то ножки 6 микросхемы К176ИА4 нужно соединить с массой, а если с общим анодом (плюс), то с плюсовым источником питания. На схеме изображен общий анод.
Я также цитирую печатную плату. На нем не сдох сам индикатор, так как они очень сильно различаются.Поэтому читателю придется уточнить плату за сам индикатор. Также обращаю внимание на то, что на плате 6 футов микросхемы подключаются по C + питанию, если у вас есть индикатор с общим «минусом», то вам нужно подключить их к питанию.
Перечень деталей:
- микросхема К176Л5 — 1 шт .;
- микросхема К176ИА4 — 2 штуки;
- резистор 1 МОм; Резистор
- 220 Ом;
- конденсатор 220 НФ.
Вот и все, схема в принципе не требует настройки.
Перечень радиоэлементов
Обозначение Тип Номинал номер Примечание Оценка Мой ноутбук IC1, IC2. Чип 2 В записной книжке IC3. Микросхема K176L5. 1 В схеме неверно В записной книжке C1. Конденсатор 0,22 мкФ. 1 В записной книжке R1 Резистор 1 МОм
1 В записной книжке R2 Резистор 220 О.
1 В записной книжке 7seg1, 7seg2. Светодиодный цифровой индикатор 2 В записной книжке S1. Переключатель 1 Высокопроизводительный анализ микросхем человеческого мозга с помощью мультинейронного патч-зажима нового поколения
Существенные изменения:
1) Одной из основных проблем является система очистки пипеток, адаптированная авторами. В соответствии с исходным протоколом CR Forest, необходим дополнительный этап для очистки остаточного детергента, приставшего к внешней поверхности наконечника пипетки с помощью ACSF, перед перемещением пипеток в записывающую камеру для попытки пластыря, однако Пэн и его коллеги пропустили этот шаг.Авторы заявляют, что есть веская практическая причина для пропуска этого шага, но данных в поддержку этой практики предоставлено мало или они отсутствуют. Они утверждают, что не было различий в качестве записи или электрофизиологических свойствах между пипетками вначале и после очистки, но эти утверждения должны быть подтверждены данными. В частности, было бы важно сообщить, как мембранный потенциал, входное сопротивление, синаптические события и параметры потенциала действия (амплитуда, ширина и др.) Изменяются со временем после исправления и повторного сопоставления.
Благодарим вас за понимание нашего обоснования отказа от дополнительной очистки. Мы согласны с тем, что эта практика и наше заявление о неизменной физиологии нейронов должны быть подтверждены дополнительными данными. Чтобы оценить возможное влияние нашего протокола очистки на качество записи и электрофизиологические свойства, мы провели дополнительные эксперименты на острых срезах мозга из моторной коры крыс. Мы зарегистрировали 81 нейрон в 12 срезах мозга от 2 животных (P21, P22) с помощью 28 пипеток с двумя последовательными циклами очистки и сравнили клеточную и синаптическую физиологию.Мы исключили 4 интернейрона, кроме того, были исключены 9 клеток с деполяризованным мембранным потенциалом, которые были зарегистрированы на свежих и очищенных пипетках с равной вероятностью (3/28 клеток со свежими пипетками, 2/28 клеток после первой очистки и 4/28 клеток после второй очистки. ), подробнее см. в разделе «Материалы и методы».
Мы построили график распределения клеточных и синаптических свойств свежих и очищенных пипеток на рисунке 3. Кроме того, мы рассчитали среднее относительное изменение и его доверительный интервал для всех запрошенных параметров.Мы обнаружили, что относительное изменение среднего значения этих параметров находится в пределах 10%. Мы также рассчитали доверительный интервал этих относительных средних изменений, который представляет собой границы статистически значимой эквивалентности. В целом, мы не нашли доказательств систематического воздействия нашего подхода к очистке на клеточную или синаптическую физиологию. Мы включили статистические результаты в качестве исходных данных на рис. 3 и соответствующим образом адаптировали раздел «Материалы и методы».
2) В статье слишком много внимания уделяется анализу связности и игнорируются его ограничения, например.г., ложные негативы. Авторы могут пожелать, вместо этого, подчеркнуть, что запись мультипатчей в настоящее время является единственным доступным методом для анализа прочности и краткосрочной пластичности моносинаптической связи.
Мы благодарим рецензента за то, что он поднял этот вопрос, и соглашаемся с тем, что существуют ограничения в отношении анализа связности с использованием мультипатч-записей. Мы включили параграф, посвященный потенциальным причинам ложноотрицательных результатов в Обсуждение.
Мы также согласны с тем, что синаптическая сила и кратковременная пластичность являются важными параметрами этих связей.Хотя парная конфигурация записи с фиксацией фиксации представляет собой оптимальный подход к анализу этих параметров, они также могут быть определены путем комбинирования записи с фиксацией фиксации с двухфотонным снятием каркаса глутамата или оптогенетической стимуляцией. Однако надежность пресинаптической стимуляции может быть ниже, чем при использовании метода патч-кламп. Мы подчеркнули важность этих параметров и технические преимущества мультипатч-подхода в соответствующем разделе «Обсуждение».
3) Важные отсутствующие экспериментальные детали включают в себя указание на возможность исправления ячеек во время записи из других ячеек, время, необходимое для проверки возможности соединения, и анализ распределения расстояний между записанными ячейками (например,g., является ли распределение ячеек по расстояниям, полученным для продления записи, таким же, как полученное изначально?).
Хотя проверка возможности подключения одновременно с установкой исправлений сэкономила бы время, мы воздержались от этого по нескольким практическим причинам, которые мы изложили в новом абзаце в разделе «Результаты». Мы также включили время, необходимое для проверки возможности подключения и измерения внутренних свойств ячеек (раздел «Результаты»).
Мы благодарим рецензента за то, что он поднял важный вопрос о том, что межсоматические расстояния могут влиять на вероятность соединения и что местоположения репатриированных ячеек следует контролировать, чтобы предотвратить возможное смещение.Как и предполагалось, мы проанализировали эффект очистки с расширением на межсоматическое расстояние в наших предыдущих экспериментах с предубикулумом крыс и обнаружили аналогичное распределение между кластерами, полученными с очисткой с расширением и без нее. Мы построили распределение расстояний на рисунке 5 — приложение к рисунку 1 и обсудили их в разделе «Результаты».
4) Авторы подчеркивают некоторые преимущества полуавтоматического подхода, но не определяют другие аспекты экспериментов с несколькими патчами, которые могут выиграть от автоматизации — например, сбор данных и онлайн-контроль качества, а также обнаружение соединения в реальном времени.Учитывая потенциал для сбора такого большого количества данных, следует рассмотреть формат данных (например, нейроданные без границ), совместное использование данных, автоматизацию обнаружения и анализа соединений.
Мы согласны с тем, что есть несколько аспектов экспериментов, которые можно автоматизировать в дальнейшем, и мы также убеждены, что увеличение объема данных требует стандартизации анализа и формата данных. Однако мы видим компромисс между экспериментальной гибкостью и автоматизацией сбора и анализа данных.Поскольку мы хотели максимизировать применимость для других групп и их конкретных вопросов в этом отчете, мы использовали коммерчески доступное программное обеспечение для сбора данных, в то время как автоматический анализ трассировки для обнаружения соединений, безусловно, важен и требует постоянных усилий. Хотя программное обеспечение Signal может также выполнять онлайн-анализ, мы считаем, что это необходимо только для экспериментов с замкнутым циклом. Мы включили новый абзац по этим вопросам в раздел «Обсуждение». Мы также поддерживаем усилия открытой науки и разработки стандартизированного формата данных для облегчения сотрудничества.Мы предоставили предложения по этой теме в разделе «Обсуждение».
5) Несмотря на то, что использование редких живых тканей человека и, особенно, для увеличения объема данных по каждому образцу является веским основанием для разработки систем с несколькими заплатками, возможно, еще не известно, будет ли этого достаточно, чтобы исследовать разницу между частные лица. Какие различия наблюдались при обсуждении различий между людьми? Типы ячеек, связи? Я бы посоветовал авторам смягчить это утверждение.
Мы понимаем озабоченность автора обзора относительно статистической мощности размеров нашей выборки для выявления значимых различий между людьми. Мы хотим подчеркнуть, что наша основная цель получения больших выборок от отдельных пациентов состоит не в том, чтобы определить эти различия между отдельными людьми, а, скорее, в том, чтобы получить возможность оценивать индивидуальную изменчивость. Мы считаем, что это очень важно, поскольку ткань получена от очень разнородной группы пациентов.Анализ данных на индивидуальном уровне может помочь нам определить инвариантные параметры, которые могут указывать на общие принципы коры головного мозга человека. С другой стороны, параметры с высокой индивидуальной вариабельностью следует анализировать с осторожностью и проводить дальнейшие исследования. Поэтому мы считаем, что получение статистически значимых наборов данных у одиноких пациентов является важным шагом для мотивации и руководства будущими исследованиями. Мы перефразировали и детализировали нашу претензию в рукописи, чтобы лучше отразить этот аспект (Аннотация; Введение; Обсуждение).
В нашем предварительном анализе мы не обнаружили значительных различий во взаимосвязи пирамидных клеток между пациентами, в то время как мы определили, что доверительный интервал различий в вероятности подключения находился в диапазоне от -5% до 9,5%. В целом, мы полагаем, что полный анализ и обсуждение потенциальных инвариантных и вариантных параметров выходят за рамки этого технического отчета, и их лучше рассмотреть в отдельной исследовательской статье. Мы добавили статистический анализ в соответствующие разделы «Результаты» и «Материалы и методы».
[Примечание редакции: до принятия были запрошены дополнительные исправления, как описано ниже.]
Рукопись была значительно улучшена, но остается одна проблема, которую необходимо решить перед принятием, как указано ниже:
Относительно новых экспериментов по повторному связыванию после обработки Alconox без промывки Alconox (подраздел «Окончательная последовательность удаления не требует дополнительных лунок, содержащих aCSF»). Они убедительны и показывают, что в тканях мозга молодых крыс в целом наблюдается небольшой кумулятивный эффект процесса очистки на последующее здоровье нейронов.Здесь необходимо прояснить два момента.
1) Ожидают ли авторы, что результаты со зрелой тканью мозга человека будут эквивалентны результатам с тканью мозга молодой крысы? Есть ли какие-либо ограничения, о которых нам следует знать в этом валидационном эксперименте.
2) Были ли когда-либо репатриированы одни и те же нейроны после очистки? Это позволит провести прямое сравнение свойств нейронов до и после очистки патч-пипетки.
Мы провели первые ревизионные эксперименты на крысах, потому что у нас редко есть человеческие ткани и мы не получали их во время ревизии.Мы также не пытались перепатчить одни и те же нейроны. Однако нам повезло, и мы дважды получали человеческую ткань за последние две недели, и теперь мы провели дополнительные эксперименты, чтобы оценить влияние очистки на электрофизиологические свойства человеческих нейронов.
Мы сравнили свойства нейронов, покрытых свежими (n = 24) или очищенными пипетками (n = 9, рисунок 3 — приложение к рисунку 1). Мы также перепрограммировали те же нейроны той же очищенной пипеткой (n = 9, рисунок 3 — приложение к рисунку 2) или другой свежей пипеткой (n = 5, рисунок 3 — приложение к рисунку 3).Мы смогли показать, что внутренние электрофизиологические свойства человеческих нейронов были и оставались схожими в разных условиях (статистические данные и тесты на рисунке 3 — исходные данные 1). Хотя мы действительно наблюдали значительное снижение входного сопротивления в клетках, повторно сопоставленных с помощью очищенной пипетки, несколько дополнительных факторов могли способствовать изменчивости в этих нейронах, например, эффект самовосстановления или прошедшее время. Эти не зависящие от очистки изменения отражаются в вариабельности, обнаруженной также в нейронах, повторно обработанных свежими пипетками (рис. 3 — приложение к рис. 3).
Поскольку мы показали, что сама пипетка не оказывает систематического воздействия на внутренние свойства, мы также рассмотрели возможность того, что внеклеточный раствор, в котором промывались пипетки, мог иметь эффект (рис. 3 — приложение к рисунку 4). Поэтому мы исправили кластеры нейронов (n = 19) и сравнили их свойства и синаптические связи (n = 7) до и после очистки других пипеток, чтобы смоделировать изменения во внеклеточном растворе после промывания.Опять же, мы обнаружили, что потенциал мембраны покоя и кинетика потенциала действия оставались очень стабильными (средняя относительная разница в пределах 2%), в то время как входное сопротивление и сопротивление доступа увеличивались. Мы также не наблюдали особой тенденции в постсинаптических амплитудах между этими двумя состояниями, которая показывала как небольшое увеличение, так и уменьшение (n = 7, Рисунок 3 — приложение к рисунку 5, Рисунок 3 — исходные данные 2). Мы соответствующим образом скорректировали разделы «Результаты» и «Материалы и методы» в рукописи.
В целом, мы смогли показать, что результаты наших экспериментов по очистке нейронов человека аналогичны тем, которые мы показали на нейронах крысы, даже когда та же самая клетка была репатриирована.Мы действительно увидели, что входное сопротивление уменьшилось в клетках, повторно обработанных с помощью очищенной пипетки, и увеличилось в клетках, зарегистрированных в ACSF после промывки. Хотя повторная синхронизация, время записи и нейронная изменчивость могут повлиять на эти параметры, мы не можем исключить эффект очистки пипетки в этом случае. Поэтому мы подчеркиваем, что эти валидационные эксперименты ограничиваются нашими вопросами настройки и исследования и что любая реализация нашей процедуры очистки другими должна быть тщательно проверена на параметры и в интересующей модели организма.Тем более, что прилипший детергент на пипетке может зависеть от множества факторов, которые необходимо учитывать и настраивать для каждой экспериментальной установки (подраздел «Окончательная последовательность изгнания не обязательно требует дополнительных лунок, содержащих aCSF»).
Кроме того, поскольку каждый патч-электрод использовался более одного раза, возможно, релевантным статистическим сравнением здесь является дисперсионный анализ повторных измерений, а не статистика популяции групп, как, по-видимому, показано на рис. 3J, K и L.
Спасибо за полезный совет. Мы выполнили повторные измерения ANOVA для 14 пипеток, с помощью которых были получены три успешных записи пирамидных клеток (свежие, 1x очистка, 2x очистка). Он не показал значительной тенденции, и мы включили результаты в Рисунок 3 — исходные данные 2. Мы также соответствующим образом адаптировали разделы «Результаты» и «Материалы и методы».
https://doi.org/10.