Физматика Ответы на вопросы Электромагнетизм. § 33. Способы индуцирования тока
Электромагнетизм. § 33. Способы индуцирования тока → номер 35
1. Почему в опытах по изучению магнитных явлений используются катушки, состоящие из большого числа витков?
Потому что ЭДС индукции, которые возникают в отдельных витках, суммируются, а это облегчает обнаружение индукционного тока.
2. Объясните причину возникновения и направление индукционного тока в опыте Фарадея с двумя вставленными друг в друга катушками
Индукционный ток через гальванометр наблюдался при замыкании или размыкании цепи, т. е. при изменении магнитного потока
Через наружную катушку. При замыкании ключа ток, протекающий по внутренней катушке, создает индукцию, направленную вверх в область наружной катушки (см. рис. 110а). Выберем направление обхода витка наружной катушки по ближайшей к нам стороне вправо. Вектор его площади направлен вверх, тогда изменение магнитного потока больше 0, а ЭДС индукции меньше 0. Это значит, что протекает противоположно направлению обхода контура (по ближайшей к нам стороне влево).
Это значит, что протекает противоположно направлению обхода контура (по ближайшей к нам стороне влево).
3. Почему в наружной катушке возникает индукционный ток при выдвигании внутренней катушки, подключенной к источнику тока? Как определяется его направление?
Магнитная индукция, которая создана внутренней катушкой в области наружной, определяется по правилу буравчика и направлена вниз. Поэтому магнитный поток через витки наружной катушки будет отрицательным. Изменение потока после выдвижения катушки положительно, ЭДС отрицательно. Это значит, что индукционный ток протекает противоположно направлению обхода (по ближайшей к нам стороне влево).
4. Объясните, почему возникает индукционный ток в катушке при вдвигании в нее магнита.
При вдвигании магнита в катушку изменяется магнитный поток (за счет изменения вектора магнитной индукции), следовательно, возникает индукционный ток.
При выдвигании внутренней катушки меняется магнитный поток, пронизывающий неподвижную катушку, что приводит к возникновению индукционного тока.
5. С одинаковым ли ускорением падает маленький полосовой магнит через вертикально стоящую катушку при замкнутой и разомкнутой обмотке катушки?
Если ток в катушке отсутствует, то магнитный поток через катушку не меняется. При замкнутой обмотке катушки, в ней возникает индукционный ток, который препятствует движению магнита.
| Электромагнитная индукция | |
1831 г. — М. Фарадей обнаружил, что в замкнутом проводящем контуре при изменении магнитного поля возникает так называемый индукционный ток. (Индукция, в данном случае, — появление, возникновение). | |
Индукционный ток в катушке возникает при перемещении постоянного магнита относительно катушки; при перемещении электромагнита относительно катушки; при перемещении сердечника относительно электромагнита, вставленного в катушку; при регулировании тока в цепи электромагнита; при замыкании и размыкании цепи | |
Появление тока в замкнутом контуре при изменении магнитного поля, пронизывающего контур, свидетельствует о действии в контуре сторонних сил (или о возникновении ЭДС индукции). Явление возникновения ЭДС в замкнутом проводящем контуре при изменении магнитного поля (потока), пронизывающего контур, называется электромагнитной индукцией. Или: явление возникновения электрического поля при изменении магнитного поля (потока), называется электромагнитной индукцией. | |
Закон электромагнитной индукции При всяком изменении магнитного потока через проводящий замкнутый контур в этом контуре возникает электрический ток. I зависит от свойств контура (сопротивление): . e не зависит от свойств контура: .ЭДС индукции в замкнутом контуре прямо пропорциональна скорости изменения магнитного потока через площадь, ограниченную этим контуром. | |
Основные применения электромагнитной индукции: генерирование тока (индукционные генераторы на всех электростанциях, динамомашины), трансформаторы. |
|
Возникновение индукционного тока — следствие закона сохранения энергии! В случае 1: При приближении магнита, увеличении тока, замыкании цепи: ; Магнитный поток Ф → ΔФ>0.Чтобы компенсировать это изменение (увеличение) внешнего поля, необходимо магнитное поле, направленное в сторону, противоположную внешнему полю: , где — т.н. индукционное магнитное поле. В случае 2: при удалении магнита, уменьшении тока, размыкании цепи: . Магнитный поток Ф → ΔФ<0. Чтобы компенсировать это изменение (уменьшение), необходимо магнитное поле, сонаправленное с внешним полем: . | |
Источником магнитного поля является ток. Поэтому: Возникающий в замкнутом контуре индукционный ток имеет такое направление, что созданный им поток магнитной индукции через площадь, ограниченную контуром, стремится компенсировать то изменение потока магнитной индукции, которое вызывает данный ток (правило Ленца). |
|
Ток в контуре имеет отрицательное направление (),еслипротивоположно (т.е. ΔΦ>0). Ток в контуре имеет положительное направление (), если совпадает с , (т.е. ΔΦ<0). Поэтому с учетом правила Ленца (знака) выражение для закона электромагнитной индукции записывается: . Данная формула справедлива для СИ (коэффициент пропорциональности равен 1). В других системах единиц коэффициент другой. | |
Если контур (например, катушка) состоит из нескольких витков, то , где n – количество витков. Все предыдущие формулы справедливы в случае линейного (равномерного) изменения магнитного потока. В произвольном случае закон записывается через производную: , где e – мгновенное значение ЭДС индукции. | |



явление электромагнитной индукции(опыты Фарадея)
явление электромагнитной индукции(опыты Фарадея)
В 1831 г. Фарадей открыл, что во всяком замкнутом проводящем контуре при изменении потока магнитной индукции через поверхность, ограниченную этим контуром, возникает электрический ток. Это явление называют электромагнитной индукцией, а возникающий ток индукционным.
Рис. 105. |
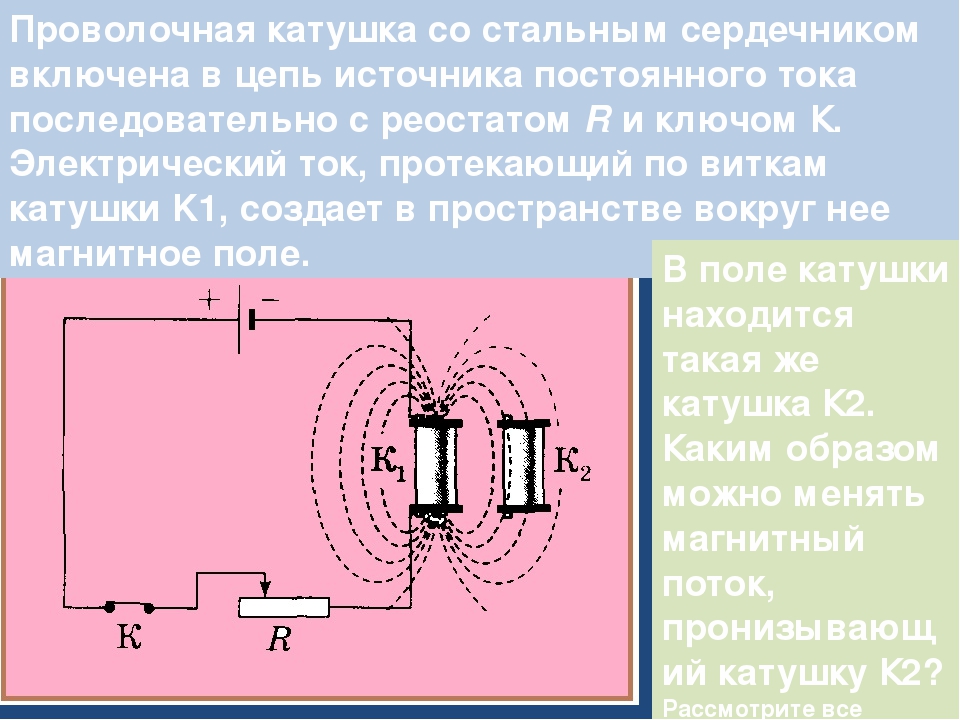
Величина индукционного тока не зависит от способа, которым вызывается изменение потока магнитной индукции Ф, но определяется лишь скоростью изменения Ф, т. е. значением dФ/dt. При изменении знака dФ/dt меняется также направление тока. Поясним сказанное следующим примером. На рис. 105 изображен контур 1, силу тока в котором i1 можно менять с помощью реостата. Ток i1 создает магнитное поле, пронизывающее контур 2. Если увеличивать ток i1 поток магнитной индукции Ф через контур 2 будет расти. Это приведет к появлению в контуре 2 индукционного тока i2, регистрируемого гальванометром. Уменьшение тока i1 обусловит убывание потока магнитной индукции через второй контур, что приведет к появлению в нем индукционного тока иного направления, чем в первом случае.
Заполнение всего пространства, в котором поле отлично от нуля, однородным магнетиком приводит, при прочих равных условиях, к увеличению индукционного тока в μ раз. Этим подтверждается то, что индукционный ток обусловлен изменением не потока вектора Н, а потока магнитной индукции.
Ленц установил правило, с помощью которого можно найти направление индукционного тока. Правило Ленца гласит, что индукционный ток всегда направлен так, чтобы противодействовать причине, его вызывающей. Если, например, изменение Ф вызвано перемещением контура, то возникает индукционный ток такого направления, что сила, действующая на него во внешнем поле, противится движению контура. При приближении контура 2 к первому контуру возникает ток i2 (рис. 105), магнитный момент которого направлен против внешнего поля (угол α между векторами рm и В равен π). Следовательно, согласно формуле (48.8) на контур 2 будет действовать сила, отталкивающая его от первого контура. При удалении контура 2 от первого контура возникает ток i2″, момент которого рm совпадает по направлению с В (α = 0), так что сила, действующая на контур 2, имеет направление к первому контуру.
При приближении контура 2 к первому контуру возникает ток i2 (рис. 105), магнитный момент которого направлен против внешнего поля (угол α между векторами рm и В равен π). Следовательно, согласно формуле (48.8) на контур 2 будет действовать сила, отталкивающая его от первого контура. При удалении контура 2 от первого контура возникает ток i2″, момент которого рm совпадает по направлению с В (α = 0), так что сила, действующая на контур 2, имеет направление к первому контуру.
Пусть контур 2 неподвижен, и ток индуцируется в нем путем изменения тока i1 в первом контуре. В этом случае индуцируется ток i2 такого направления, что создаваемый им собственный магнитный поток стремится ослабить изменения внешнего потока, приведшие к появлению индукционного тока. При увеличении i1 т. е. возрастании внешнего магнитного потока, направленного вправо, возникнет ток i2”, создающий поток, направленный влево. При уменьшении i1 возникает ток I, собственный магнитный поток которого направлен так же, как и внешний поток, и, следовательно, стремится поддержать внешний поток неизменным.
ЯВЛЕНИЕ ЭЛЕКТРО-МАГ. ИНДУКЦИИ. ПРАВИЛО ЛЕНЦА.
Явление электромагнитной индукции заключается в том, что в замкнутом проводящем контуре при изменении потока магнитной индукции, охватываемой этим контуром возникает электрический ток, называемый индукционным потоком. Эксперементально установленно, что величина индукции тока не зависит от способов изменения магнитного потока, а определяет лишь скорость изменения. Индукционный ток в проводнике может возникнуть только под действием ЭДС. ЭДС, возникшая в проводнике при изменении магнитного потока, называется ЭДС-индукции. Согласно закону Фарадея: ε инд=k dФm/dt. Направление индукции тока определяется по правилу Ленца. При всяком изменении магнитного потока сквозь поверхность натянутую на замкнутый контур, в нем возникает индукционный ток такого направления, что его магнитное поле препятствует изменению магнитного потока. С учетом правила Ленца закон Фарадея имеет вид: ε инд = — dФm/dt; Поскольку для замкнутого контура dФm=dψ, то ε инд = — dψ/dt
Электромагнитная индукция — презентация онлайн
Prezentacii. com
comЭлектромагнитная
индукция
Сравнение электростатического и
магнитного полей
Электростати- магнитное
ческое
Источник поля
Что служит индикатором поля?
Характеристика поля
Линии поля: замкнуты или
незамкнуты
Характер поля ( потенциальное
или вихревое)
Знаем:
o Электрическое поле создается
неподвижными заряженными
частицами
o Магнитное поле – движущимися, т.е.
электрическим током
Умеем:
o Превращать электричество в магнетизм
Задача:
«Превратить
магнетизм в
электричество»
1821-1831 годы
М. Фарадей
Благодаря этому открытию были сконструированы
устройства: генераторы, трансформаторы и т.д.
Майкл Фарадей (1791 — 1867)
Вопросы к данному
эксперименту:
1. Что наблюдаем в данном
эксперименте?
2. Что является причиной появления
тока в катушке?
Электрический ток,
возникший в контуре,
будем называть
индукционным.

А явление возникновения
тока при данных
условиях, -явлением
электромагнитной
индукции
o Явление электромагнитной индукции
заключается в возникновении
электрического тока в проводящем контуре,
при изменении числа линий магнитной
индукции, пронизывающих этот контур (при
изменении магнитного потока ).
Способы индуцирования тока
(Опыты Фарадея)
Магнитный поток, пронизывающий
катушку, может изменяться по трем
причинам:
o за счет изменения магнитного поля, в
котором находится неподвижная
катушка;
o за счет движения самой
катушки в магнитном поле
o Если по катушке идет
переменный ток
Определение явления ЭМИ
o Явление электромагнитной индукции
заключается в возникновении
электрического тока в проводящем
контуре, который либо покоится в
переменном во времени магнитном
поле, либо движется в постоянном
магнитном поле, таким образом, что
меняется магнитный поток,
пронизывающий этот контур.

Направление тока
Правило Ленца.
индукционный ток всегда имеет
такое направление, что
создаваемое им магнитное поле
препятствует изменению
магнитного потока, вызывающего
индукционный ток.
Применение правила Ленца:
ü Установить направление линий магнитной
индукции В внешнего поля
ü Выяснить , увеличивается или уменьшается
магнитный поток
ü Установить направление линий магнитной
индукции В’ магнитного поля индукционного тока.
при ∆Ф>0, В’ ↑ ↓ В
при ∆Ф
ü Зная направление линий магнитной индукции В’,
найти направление индукционного тока,
пользуясь правилом буравчика или правилом
правой руки
Правило Ленца
Пример 1
По правилу буравчика: B’
при ∆Ф>0, В’ ↑ ↓ В
Направление B
Определяем полюса магнита
Самостоятельная работа
o 1 вариант
Определить полюса
магнита.
o 2 вариант
Определить направление
движения магнита
ЭДС индукции
Ø В цепи появляется электрический ток
в том случае, когда на свободные
заряды действуют сторонние силы.

Ø При изменении магнитного потока
через контур, в контуре появляются
сторонние силы, действие которых
характеризуется ЭДС, называемой
ЭДС индукцией
o Э
Д
С
и
н
д
у
к
ц
и
и
в
з
а
м
к
Закон электромагнитной
индукции
o Сила индукционного тока пропорциональна
скорости изменения магнитного потока через
поверхность, ограниченную контуром:
o
ЭДС в проводнике,
движущемся в магнитном
поле
При движении проводника его свободные заряды
движутся вместе с ним. Поэтому на заряды со стороны
магнитного поля действует сила Лоренца. Она-то и
вызывает перемещение зарядов внутри проводника.
Следовательно, ЭДС индукции в движущемся
проводнике имеет магнитное происхождение.
o
ЭДС индукции в неподвижном
проводнике.
o На неподвижные заряды может оказывать
действие только электрическое поле. Но
индукционный ток появляется в результате
действия переменного магнитного поля.

Это заставляет предположить, что
электроны в неподвижном проводнике
приводятся в движение электрическим
полем, которое порождается переменным
магнитным полем
Теория Максвелла
o
Изменяясь во
времени, магнитное
поле порождает
электрическое
поле
свойства вихревого
электрического поля
o Источник поля: изменяющее магнитное поле
o Индикатором поля являются электрические
заряды
o Силовые линии представляют собой
замкнутые линии. Поле носит вихревой
характер.
o Работа вихревого поля на замкнутом пути не
равна нулю.
o Характеристика поля: напряженность:
F= q E
Явление ЭМИ в новом свете:
o Явление электромагнитной индукции
заключается в возникновении
вихревого электрического поля,
вызывающего электрический ток в
замкнутом контуре, при изменении
магнитного потока пронизывающего
этот контур.
Опыты Генри
o Если по катушке идет переменный ток,
то магнитный поток, пронизывающий
катушку, меняется.
 Поэтому возникает
Поэтому возникаетЭДС в том же самом проводнике, по
которому идет переменный ток.
o Это явление называется самоиндукцией:
Возникновение ЭДС индукции в
проводящем контуре при изменении
в нем силы тока.
Всего хорошего!
До новых встреч!
Самоиндукция, индуктивность. самоиндукция каждый проводник, по которому протекает эл.ток, находится в собственном магнитном поле. урок явление самоиндукции
Урок физики № 47 в 9 классе.
Дата:
Тема: «Самоиндукция»
Цель урока :
- Изучение сущности явления самоиндукции; знакомство с величиной индуктивность, формулой для расчета энергии магнитного поля, выяснение физического смысла этой формулы.
- Развитие логического мышления, внимания, умений анализировать результаты эксперимента, делать выводы.
- Воспитание культуры умственного труда; интереса к физике; формирование коммуникативных качеств личности.
Тип урока: комбинированный.
Форма урока: смешанная.
Д/З: § 49, 50.
Ход урока
- Орг. момент.
- Проверка д/з.
- Устный опрос.
- Явление электромагнитной индукции.
- Способы индуцирования тока.
- Индивидуальная работа по карточкам.
- Объяснение нового материала.
- Дополнительный материал.
Направление индукционного тока.
Вопросы к учащимся для актуализации прежних знаний:
- Назвать две серии опытов Фарадея по исследованию явления электромагнитной индукции (возникновение индукционного тока в катушке при вдвигании и выдвигании магнита или катушки с током; возникновение индукционного тока в одной катушке при изменении тока в другой путем замыкания-размыкания цепи или использования реостата).
- Зависит ли направление отклонения стрелки гальванометра от направления движения магнита относительно катушки? (зависит: при приближении магнита к катушке стрелка отклоняется в одну сторону, при удалении магнита — в другую).
- Чем отличается (судя по показаниям гальванометра) индукционный ток, возникающий в катушке при приближении магнита, от тока, возникающего при удалении магнита (при одинаковой скорости движения магнита)? (ток отличается направлением).
Таким образом, при движении магнита относительно катушки направление отклонения стрелки гальванометра (а, значит, и направление тока) может быть различным. Сформулируем при помощи опыта Ленца правило нахождения направления индукционного тока (видеоролик «Демонстрация явления электромагнитной индукции»).
Объяснение опыта Ленца: Если приближать магнит к проводящему кольцу, то оно начнет отталкиваться от магнита. Это отталкивание можно объяснить только тем, что в кольце возникает индукционный ток, обусловленный возрастанием магнитного потока через кольцо, а кольцо с током взаимодействует с магнитом.
Правило Ленца и закон сохранения энергии.
возрастает , то направление индукционного тока в контуре таково, что вектор магнитной индукции созданного этим током поля направлен противоположно вектору магнитной индукции внешнего магнитного поля.
Если магнитный поток через контур уменьшается , то направление индукционного тока таково, что вектор магнитной индукции созданного этим током поля сонаправлен вектору магнитной индукции внешнего поля.
Формулировка правила Ленца: индукционный ток имеет такое направление, что созданный им магнитный поток всегда стремится скомпенсировать то изменение магнитного потока, которое вызвало данный ток.
Правило Ленца является следствием закона сохранения энергии.
- Явление самоиндукции.
- Прежде, чем рассмотреть явление самоиндукции, вспомним, в чем заключается суть явления электромагнитной индукции — это возникновение индукционного тока в замкнутом контуре при изменении магнитного потока, пронизывающего этот контур. Рассмотрим один из вариантов опытов Фарадея: Если в цепи, содержащей замкнутый контур (катушку) менять силу тока, то в самом контуре возникнет ещё и индукционный ток. Этот ток также будет подчиняться правилу Ленца.
Рассмотрим опыт по замыканию цепи, содержащей катушку. При замыкании цепи с катушкой определенное значение силы тока устанавливается лишь спустя некоторое время.
- Видеофрагмент «Самоиндукция»
- Определение самоиндукции: САМОИНДУКЦИЯ — возникновение вихревого электрического поля в проводящем контуре при изменении силы тока в нем; частный случай электромагнитной индукции
.
Вследствие самоиндукции замкнутый контур обладает «инертностью»: силу тока в контуре, содержащем катушку, нельзя изменить мгновенно.
3. Индуктивность.
Ф=LI
Единицы измерения индуктивности в системе СИ: [L] = 1 = 1 Гн (генри).
- Применение и учет самоиндукции в технике .
Вследствие явления самоиндукции при размыкании цепей, содержащих катушки со стальными сердечниками (электромагниты, двигатели, трансформаторы) создается значительная ЭДС самоиндукции и может возникнуть искрение или даже дуговой разряд. В качестве домашнего задания предлагаю (по желанию) подготовить презентацию на тему «Как устранить нежелательную самоиндукцию при размыкании цепи?».
- Энергия магнитного поля
- Закрепление.
- Упр. 41 — устно.
- № 830, 837 — у доски.
- № 834 — на рабочих местах.
- Рефлексия.
- Итог урока.
- Д/з.
style=»&6�#:.��I �E s New Roman»»> Опыт Фарадея.
Магнитные и электрические поля связаны друг с другом. Эл. ток способен вызывать появление магнитного поля. А не может ли магнитное поле создать электрический ток? Эту задачу пытались решить многие ученые в начале 19 века. Но первый решающий вклад в открытии ЭМ взаимодействий был сделан Майклом Фарадеем.
“Превратить магнетизм в электричество”- записал Фарадей в своем дневнике. 1821г. И только через10 лет он смог решить эту задачу. Мы с вами откроем, то, что Фарадей не мог открыть 10 лет, за несколько минут. Фарадей не мог понять одного: что только движущийся магнит вызывает ток. Покоящийся магнит не вызывает в ней тока. Какие же опыты проводил Фарадей? Давайте повторим опыты Фарадея, с помощью которых он открыл явление ЭМИ.
Демонстрация: возникновение индукционного тока (катушка, миллиамперметр, постоянный магнит)
Определение: Возникновение в замкнутом проводнике электрического тока, обусловленное изменение магнитного поля называют явлением ЭЛЕКТРОМАГНИТНОЙ ИНДУКЦИИ.
Полученный ток называют — индукционным.
ВЫВОД: Индукционный ток возникает только при относительном перемещении катушки и магнита. Направление индукционного тока зависит от направления вектора В внешнего магнитного поля.
- Способы получения индукционного тока.
Индукционный ток в замкнутом контуре появляется только в том случае, когда изменяется магнитный поток, который проходит через площадь охваченную контуром.
Работа в группах (использование учебника, Интернета)
1 группа: 1 способ (рис. 127)
- Закрепление нового материала.
- Упр. 39 (1,2) — устно;
- Упр. 40 (2) — устно.
- Рефлексия.
- Итог урока.
- Д/з.
Тема урока : САМОИНДУКЦИЯ .
Цели урока :
Обучающая : ознакомить уч-ся с явлением самоиндукции, сформировать знания по данному явлению.
Развивающая: активизировать мышление школьников, развивать мотивацию изучения физики.
Воспитательная : воспитывать интерес к предмету.
Ход урока:
Тип урока : комбинированный.
I Организационная часть.
II Этап постановки целей и задач урока: на данном уроке мы узнаем, как и кем было открыто явление самоиндукции, рассмотрим опыт, с помощью которого продемонстрируем это явление, определим, что самоиндукция — это частный случай электромагнитной индукции. В конце урока введем физическую величину, показывающую зависимость ЭДС самоиндукции от размеров и форм проводника и от среды, в которой находится проводник, т.е. индуктивность.
III Этап актуализации опорных знаний:
Вопросы классу:
1. Как формулируется закон эл.магнитной индукции.?
2. Записать закон эл. магнитной индукции?
3.Что означает знак «- «?
4. Почему закон эл.магнитной индукции формулируется для ЭДС,а не для тока7
5. Какое поле называют » вихревым»?
6.Что такое токи Фуко?
IV Этап изучения нового материала:
Самоиндукция
а. Биографические сведения об ученом открывшем явление
Основы электродинамики были заложены Ампером в 1820 году. Работы Ампера вдохновили многих инженеров на конструирование различных технических устройств, таких как электродвигатель (конструктор Б.С. Якоби), телеграф (С. Морзе), электромагнит, конструированием которого занимался известный американский ученый Генри.
Джозеф Генри (рис. 1) прославился благодаря созданию серии уникальных мощнейших электромагнитов с подъемной силой от 30 до 1500 кг при собственной массе магнита 10 кг. Создавая различные электромагниты, в 1832 году ученый открыл новое явление в электромагнетизме – явление самоиндукции. Именно этому явлению посвящен данный урок.
Рис. 1. Джозеф Генри
Джозеф Генри -1832г.
б. Демонстрация схемы цепи:
Генри изобретал плоские катушки из полосовой меди, с помощью которых добивался силовых эффектов, выраженных более ярко, чем при использовании проволочных соленоидов. Ученый заметил, что при нахождении в цепи мощной катушки ток в этой цепи достигает своего максимального значения гораздо медленнее, чем без катушки.
Рис. 2. Схема экспериментальной установки Д. Генри
Рис. 3. Различный накал лампочек в момент включения цепи
При замыкании ключа первая лампа вспыхивает практически сразу, вторая — с заметным опозданием.
ЭДС индукции в цепи этой лампы велика, и сила тока не сразу достигает своего значения.
При размыкании ключа ток в цепи уменьшается ЭДС индукции в цепи мало, а индукционный ток направлен в ту же сторону, что и собственный ток витка. Это приводит к замедлению убывания собственного тока -вторая лампа гаснет не сразу.
Вывод: при изменении тока в проводнике возникает электромагнитная индукция в том же проводнике, что порождает индукционный ток, направленный таким образом, чтобы препятствовать любому изменению собственного тока в проводнике. В этом заключается явление самоиндукции. Самоиндукция- это частный случай электромагнитной индукции. Формулы для нахождения потока магнитной индукции и ЭДС самоиндукции.
Основные выводы: Самоиндукция- это явление возникновения электромагнитной индукции в проводнике при изменении силы тока, протекающего сквозь этот проводник.
Электродвижущая сила индукции прямо пропорциональна скорости изменения тока, протекающего сквозь проводник, взятого со знаком минус. Коэффициент пропорциональности называют индуктивностью , которая зависит от геометрических параметров проводника:
Проводник имеет индуктивность, равную 1 Гн, если при скорости изменения тока в проводнике, равной 1 А в секунду, в этом проводнике возникает электродвижущая сила самоиндукции, равной 1В.
С явлением самоиндукции человек сталкивается ежедневно. Каждый раз, включая или выключая свет, мы тем самым замыкаем или размыкаем цепь, при этом возбуждая индукционные токи. Иногда эти токи могут достигать таких больших величин, что внутри выключателя проскакивает искра, которую мы можем увидеть.
Просмотр фрагмента диска » Самоиндукция в быту и технике «
V Этап закрепления нового материала.
Вопросы классу:
1. Что называют самоиндукцией?
2. Как направлены по отношению к току линии напряженности вихревого электрического поля в проводнике при увеличении и уменьшении силы тока?
3. Что называют индуктивностью?
4. Что принимают за единицей индуктивности?
5. Чему равна ЭДС самоиндукции?
Решение задач: Марон, стр. 23 В1. Рымкевич № 931, 932, 934, 935, 926.
VI Домашнее задание : п. 15, упр. Марон, стр.102 (1-й В 1-6)
Проявление явления самоиндукции Замыкание цепиРазмыкание цепи При замыкании в эл.цепи нарастает ток, что вызывает в катушке увеличение магнитного потока, возникает вихревое эл.поле, направленное против тока, т.е. в катушке возникает ЭДС самоиндукции, препятствующая нарастанию тока в цепи (вихревое поле тормозит электроны). В результате Л1 загорается позже, чем Л2. При размыкании эл.цепи ток убывает, возникает уменьшение м.потока в катушке, возникает вихревое эл.поле, направленное как ток (стремящееся сохранить прежнюю силу тока), т.е. в катушке возникает ЭДС самоиндукции, поддерживающая ток в цепи. В результате Л при выключении ярко вспыхивает.
ИНДУКТИВНОСТЬ От чего зависит ЭДС самоиндукции? Эл.ток создает собственное магнитное поле. Магнитный поток через контур пропорционален индукции магнитного поля (Ф ~ B), индукция пропорциональна силе тока в проводнике (B ~ I), следовательно магнитный поток пропорционален силе тока (Ф ~ I). ЭДС самоиндукции зависит от скорости изменения силы тока в эл.цепи, от свойств проводника (размеров и формы) и от относительной магнитной проницаемости среды, в которой находится проводник. Физическая величина, показывающая зависимость ЭДС самоиндукции от размеров и формы проводника и от среды, в которой находится проводник, называется коэффициентом самоиндукции или индуктивностью.
ЭНЕРГИЯ МАГНИТНОГО ПОЛЯ ТОКА Вокруг проводника с током существует магнитное поле, которое обладает энергией. Откуда она берется? Источник тока, включенный в эл.цепь, обладает запасом энергии. В момент замыкания эл.цепи источник тока расходует часть своей энергии на преодоление действия возникающей ЭДС самоиндукции. Эта часть энергии, называемая собственной энергией тока, и идет на образование магнитного поля. Энергия магнитного поля равна собственной энергии тока. Собственная энергия тока численно равна работе, которую должен совершить источник тока для преодоления ЭДС самоиндукции, чтобы создать ток в цепи.
Энергия магнитного поля, созданного током, прямо пропорциональна квадрату силы тока. Куда пропадает энергия магнитного поля после прекращения тока? — выделяется (при размыкании цепи с достаточно большой силой тока возможно возникновение искры или дуги)
Согласно правилу Ленца, индуктивный ток, который возникает в замкнутом контуре, всегда противодействует изменению внешнего магнитного потока, которое вызвала его появление. Сегодня мы будем рассматривать случай, когда появление электромагнитной индукции обусловлено изменение силы тока, походящего через катушку с большим количеством витков. Если причина возникновения индукционного тока состоит в возрастании тока, то индукционный ток своим магнитным полем будет противодействовать этому возрастанию. Убедиться в этом можно на следующем опыте. Соединим параллельно две лампочки, к первой лампочке ток попадает, проходя через реостат, а ко второй лампочке, проходя через катушку индуктивности, причем число витков в этой катушке достаточно велико, а внутри находится сердечник, состоящий из соединенных между собой пластин трансформаторной стали (магнитное поле, которое будет возникать вокруг такой катушки, велико). Замкнем ключом цепь. Обе лампочки загорелись, но вторая лампочка, загорелась с видимым запозданием. В чем причина данного явления? В момент замыкания ключа, общая сила тока I, и силы тока в каждой ветви I1 и I2 начинают возрастать. А если вокруг проводников возникает усиление магнитного поля, тогда, в соответствии с правилом Ленца, в реостате и катушке возникают индукционные токи, которые будут препятствовать своим действием дальнейшему возрастанию силы тока в цепи. Конечно же, магнитное поле, которое возникнет вокруг катушки с током, более сильное, поэтому лампочка номер два загорается позже.
Обратите внимание, что в опытах, которые мы рассматривали ранее, индукционный ток в контуре возникал вследствие воздействия внешнего магнитного поля. В нашем примере, индукционный ток в цепи возник, по причине изменения силы тока в цепи. Это явление получило название явление самоиндукции. Явление самоиндукции — это явление, обусловленное возникновением индукционного тока в проводнике или катушке, вследствие изменения тока в ней. Возникший ток называют током самоиндукции. Лампочка загорелась позже, проходя через катушку, т.к. в катушке индукционный ток больше, чем в реостате (катушка имеет большее число витков и сердечник). Поэтому говорят, что она обладает большей индуктивностью, чем реостат.
Что же такое индуктивность? Индуктивность — это новая физическая величина, с помощью которой можно оценить способность катушки противодействовать изменению силы тока в ней. Обозначают индуктивность буквой L (эль). Единицы изменения индуктивности в международной системе единиц (СИ) — генри (Гн). Индуктивность различных катушек будет различной. Она зависит от размеров и формы катушки, числа витков, наличие сердечника и материала, из которого он изготовлен. И конечно, чем большей индуктивностью обладает катушка, тем с большим запозданием будет загораться лампочка.
Проведем второй опыт, который продемонстрирует явление самоиндукции при размыкании цепи. В схеме, которую мы собирали ранее, проведем некоторые замены. Уберем первую лампочку, а к катушке параллельно подсоединим неоновую лампочку, которую на схеме обозначим Лн (эл с индексом эн). При замыкании цепи, мы наблюдаем горение только одной лампочки. Напряжение на источнике тока меньше, чем необходимо для горения неоновой лампочки (напряжение должно быть не менее 80 Вольт). Разомкнем цепь, лампочка накаливания потухает, а неоновая лампочка озаряется кратковременной вспышкой.
Почему так происходит? При уменьшении тока в цепи в катушке возникает индукционный ток, своим магнитным полем, препятствующий уменьшению тока в цепи. Причем возникающий индукционный ток настолько велик, что его напряжения достаточно для горения неоновой лампочки, но он очень быстро ослабевает.
Подумайте и ответьте на вопрос, в каком случае в цепи возникает явление самоиндукции?
А) при уменьшении тока в цепи,
Б) при возрастании тока в цепи,
В) в обоих случаях.
Явление самоиндукции возникает при прохождении через катушку переменного тока (это может быть и увеличение тока и уменьшение).
При замыкании цепи индукционный ток
А) препятствует увеличению тока в цепи,
Б) способствует увеличению тока в цепи,
В) не влияет на протекание тока в цепи.
При замыкании ключа, возникающий индукционный ток препятствует возрастанию тока в цепи. Самоиндукция возникает во всех проводниках, при изменении силы тока в цепи, однако он будет заметен и окажет существенное влияние на другие элементы в цепи, только в случае использования катушки с достаточно большим количеством витков и сердечником.
Электромагнитная индукция. Самоиндукция. Взаимоиндукция
«При всяком изменении магнитного потока в цепи индуцируется э.д.с.»(закон электромагнитной индукции Фарадея).
Необходимо выполнение одного из двух условий:
· проводник должен перемещаться в магнитном поле или
· магнитное поле должно перемещаться вокруг проводника.
1. Как движение электрона создаёт магнитное поле, так и магнитное поле, пересекая проводник, вызывает направленное движение свободных электронов в проводнике, т.е. ток.
Ни один вид энергии не может быть получен без затраты какого-либо другого вида энергии. Так, если магнит лежит около проводника, то в проводнике никакого тока не будет. Он возникает только при перемещении магнита около проводника или проводника около магнита. В этом случае механическая энергия превращается в электрическую.
Индуцированная э.д.с. прямо пропорциональна индукции магнитного поля В, длине проводника ℓ, скорости его перемещения V в направлении, перпендикулярном силовым линиям поля. е = В∙ℓ∙V.
Рис. 2.11. Индуцирование э.д.с. в проводнике
при перемещении его в магнитном поле
Если проводник движется под углом α к направлению поля, то
е =В∙ℓ∙V∙sinα.
Направление индуцированной э.д.с. определяется по правилу правой руки(рис.2.12):
«Если ладонь правой руки расположить так, чтобы силовые линии поля входили в ладонь, большой отогнутый палец совместить с направлением движения проводника, то вытянутые четыре пальца укажут направление индуцированной э.д.с.».
Возникновение э.д.с. объясняется действием сил магнитного поля на находящиеся в проводниках свободные электроны, которые начинают двигаться вдоль проводника.
В результате этого движения на одной стороне проводника накопятся свободные электроны и возникнет отрицательный электрический заряд.
Рисунок 2.12.
Правило правой руки
На другом конце ввиду недостатка электронов появится положительный заряд. Разность потенциалов на концах проводника численно равна индуцированной в проводнике э.д.с.
Индуцирование э.д.с. в проводнике происходит независимо от того, включён он в электрическую цепь или нет. Если присоединить концы этого проводника к приёмнику электрической энергии, то под влиянием разности потенциалов на концах проводника по замкнутой цепи потечёт электрический ток. Если проводник перемещается вдоль силовых линий, т.е. как бы скользит по ним, то э.д.с. в нём не возникает.
2. Если каким-либо образом изменять магнитный поток, пронизывающий неподвижный виток, или перемещать само поле, то индуцированная э.д.с.
e =
Направление э.д.с. в неподвижном замкнутом контуре определяется по закону Максвелла:
Э.д.с., индуцированная в замкнутом контуре, равна скорости изменения магнитного потока ΔΦ, пронизывающего этот контур. Δt–время, в течение которого происходит изменение потока.
Иными словами, чем быстрее изменяется магнитный поток, пронизывающий замкнутый контур, тем больше индуцированная э.д.с.
Правило Ленца:
«Направление индуктированного тока всегда таково, что он противодействует причине, вызвавшей его появление»(например, движению проводника).
На принципе явления электромагнитной индукции основано устройство электрических генераторов, двигателей и трансформаторов. Для индуцирования э.д.с. в них применяется три способа:
Изменение тока в катушке 1 (рис.2.13), в магнитном поле которой расположена вторая катушка 2. При этом непрерывно изменяется магнитный поток, охватываемый второй катушкой, и в ней, а также и в первой катушке, будут индуцироваться электродвижущие силы е2и е1. Этот способ используют в трансформаторах.
Рис. 2.13. Индуцирование э.д.с. в трансформаторах
Вращение магнитного поля, созданного постоянными магнитами или электромагнитами 3, относительно неподвижных катушек 4 (рис.2.14). При этом непрерывно изменяется магнитный поток, пронизывающий каждую катушку, и в них индуцируется э.д.с. е. Такой способ используют в основном в машинах переменного тока.
Рис. 2.14. Индуцирование э.д.с.
в машинах переменного тока
Вращение витков 6 или катушек в постоянном магнитном поле, созданном неподвижными постоянными магнитами 5 или электромагнитами (рис. 2.15). При этом непрерывно изменяется магнитный поток, охватываемый каждым витком или катушкой, вследствие чего в них индуцируется э.д.с. е. Этот способ используют в машинах постоянного тока.
Рис. 2.15. Индуцирование э.д.с.
в машинах постоянного тока
Э.д.с. самоиндукции.
Если по витку протекает ток, изменяющийся по величине или направлению, то в нем наводится э.д.с., которая условно называется э.д.с. самоиндукции.
Направление э.д.с. самоиндукции препятствует изменению вызвавшего ее тока.
При постоянном токе этот процесс наблюдается в момент замыкания и размыкания цепи. В момент замыкания магнитный поток, создаваемый протекающим по цепи током, увеличивается, а появляющаяся э.д.с. препятствует увеличению тока. В момент размыкания ток уменьшается, а э.д.с. самоиндукции препятствует уменьшению тока. Т.е. при замыкании и размыкании цепей ток нарастает и падает постепенно.
Если замкнутый проводник состоит из одного витка, то магнитный поток, пронизывающий контур этого проводника при постоянной магнитной проницаемости пропорционален току, протекающему по проводнику.
Э.д.с. самоиндукции пропорциональна скорости изменения тока.
Рисунок 2.16.
Э.д.с. самоиндукции
Особенно сильно проявляет себя э.д.с. самоиндукции при размыкании цепей, содержащих катушки с большим числом витков W и стальными сердечниками (обмотки электродвигателей). В этом случае э.д.с. самоиндукции может во много раз превышать напряжение источника и, суммируясь с ним, служить причиной возникновения перенапряжений и искрения или дуги. Поэтому в таких устройствах предусматривают дугогасительные устройства.
Э.д.с. взаимоиндукции.
Взаимоиндукцией называется явление индуцирования э.д.с. в проводнике или катушке при изменении магнитного потока, создаваемого другим проводником или катушкой. Индуцируемая таким образом э.д.с. называется э.д.с взаимоиндукции.
Рисунок 2.17.
Э.д.с. взаимоиндукции
Э.д.с. взаимоиндукции, как и самоиндукции, пропорциональна скорости изменения тока, создающего этого поле, кроме того зависит от числа витков обеих катушек и их взаимного расположения.
Взаимоиндукция дает возможность связывать посредством магнитного поля различные электрические цепи. Явление взаимоиндукции широко используются в трансформаторах, радиотехнических устройствах и устройствах автоматики. Однако в некоторых случаях возникновение э.д.с. взаимоиндукции является нежелательным.
Например, э.д.с. взаимоиндукции, индуцированные в линиях связи (телефонных и телеграфных проводах), проложенных вдоль высоковольтных линий электропередачи или вдоль контактной сети электрифицированных железных дорог переменного тока, создают помехи при передаче телефонных или телеграфных сигналов. Поэтому линии связи стремятся располагать перпендикулярно проводам линий электропередачи или выполнять их в виде кабельных линий, защищенных металлическими экранами.
Контрольные вопросы
1. Какой способ применяют для индуцирования э.д.с. в машинах переменного тока?
2. Дайте определение э.д.с. самоиндукции?
3. Что представляют собой вихревые токи?
4. Какой способ применяют для индуцирования э.д.с. в машинах постоянного тока?
5. Дайте определение явления электромагнитной индукции.
6. Объясните причины возникновения э.д.с.
7. Сформулируйте правило правой руки.
8. Сформулируйте правило Ленца.
9. Перечислите способы индуцирования э.д.с. в генераторах, двигателях и трансформаторах.
10. Дайте определение э.д.с. взаимоиндукции?
11. Как определяется направление индуцированной э.д.с.?
В чем суть явления электромагнитной индукции
В 1821 году Майкл Фарадей записал в своем дневнике: «Превратить магнетизм в электричество». Через 10 лет эта задача была им решена. В 1831 г. Майкл Фарадей установил, что электрический ток в контуре может возникать не только при движении проводника в магнитном поле, но и при любом изменении магнитного потока (рис. 3.13.).
Рис. 3.13. Опыты Фарадея
Электромагнитная индукция – физическое явление, заключающееся в возникновении электрического тока в замкнутом контуре при изменении магнитного потока через поверхность, ограниченную этим контуром.
Электрический ток, возникающий при явлении электромагнитной индукции называется индукционным.Индукционный ток возникает всегда, когда происходит изменение сцепленного с контуром потока магнитной индукции. Сила индукционного тока не зависит от способа изменения потока магнитной индукции, а определяется лишь скоростью его изменения.
Закон Фарадея: сила индукционного тока, возникающего в замкнутом проводящем контуре (ЭДС индукции, возникающая в проводнике), пропорциональна скорости изменения магнитного потока, сцепленного с контуром (проникающего через поверхность, ограниченную контуром), и не зависит от способа изменения магнитного потока.
Правило Ленца: индукционный ток направлен таким образом, что собственным магнитным полем препятствует изменению внешнего магнитного потока, пересекающего поверхность контура (рис. 3.14).
Рис. 3.14. Иллюстрация правила Ленца
Мгновенное значение ЭДС индукции (закон Фарадея-Ленца)
,
где – потокосцепление замкнутого проводящего контура.
Открытие явления электромагнитной индукции:
1. показало взаимосвязь между электрическим и магнитным полем;
2. предложило способ получения электрического тока с помощью магнитного поля.
Таким образом, возникновение ЭДС индукции возможно и в случае неподвижного контура, находящегося в переменном магнитном поле. Однако сила Лоренца на неподвижные заряды не действует, поэтому с ее помощью нельзя объяснить возникновение ЭДС индукции.
Опыт показывает, что ЭДС индукции не зависит от рода вещества проводника, от состояния проводника, в частности от его температуры, которая может быть даже неодинаковой вдоль проводника. Следовательно, сторонние силы с изменением свойств проводника в магнитном поле, а обусловлены самим магнитным полем.
Максвелл для объяснения ЭДС индукции в неподвижных проводниках предположил, что переменное магнитное поле возбуждает в окружающем пространстве вихревое электрическое поле, которое и является причиной возникновения индукционного тока в проводнике. Вихревое электрическое поле не является электростатическим (т. е. потенциальным).
Разность потенциалов на концах проводника, движущегося со скоростью в магнитном поле,
,
где ‑ длина проводника; ‑ угол между векторами и.
Заряд, протекающий по замкнутому контуру при изменении магнитного потока, пронизывающего этот контур,
, или ,
где R ‑ сопротивление контура.
Не нашли то, что искали? Воспользуйтесь поиском:
Лучшие изречения: Студент – человек, постоянно откладывающий неизбежность. 10611 – | 7337 – или читать все.
91.146.8.87 © studopedia.ru Не является автором материалов, которые размещены. Но предоставляет возможность бесплатного использования. Есть нарушение авторского права? Напишите нам | Обратная связь.
Отключите adBlock!
и обновите страницу (F5)
очень нужно
Магнитное взаимодействие движущихся электрических зарядов согласно представлениям теории поля объясняется следующим образом: всякий движущийся электрический заряд создает в окружающем пространстве магнитное поле, способное действовать на другие движущиеся электрические заряды.
В – физическая величина, являющаяся силовой характеристикой магнитного поля. Она называется магнитной индукцией (или индукцией магнитного поля).
Магнитная индукция – векторная величина. Модуль вектора магнитной индукции равен отношению максимального значения силы Ампера, действующей на прямой проводник с током, к силе тока в проводнике и его длине:
Единица магнитной индукции. В Международной системе единиц за единицу магнитной индукции принята индукция такого магнитного поля, в котором на каждый метр длины проводника при силе тока 1 А действует максимальная сила Ампера 1 Н. Эта единица называется тесла (сокращенно: Тл), в честь выдающегося югославского физика Н. Тесла:
Движение проводника с током в магнитном поле показывает, что магнитное поле действует на движущиеся электрические заряды. На проводник действует сила Ампера FА = IBlsin a , а сила Лоренца действует на движущийся заряд:
где a – угол между векторами B и v .
Движение заряженных частиц в магнитном поле. В однородном магнитном поле на заряженную частицу, движущуюся со скоростью перпендикулярно линиям индукции магнитного поля, действует силам , постоянная по модулю и направленная перпендикулярно вектору скорости.Под действием магнитной силы частица приобретает ускорение, модуль которого равен:
В однородном магнитном поле эта частица движется по окружности. Радиус кривизны траектории, по которой движется частица, определяется из условияоткуда следует,
Радиус кривизны траектории является величиной постоянной, поскольку сила, перпендикулярная вектору скорости, меняется только ее направление, но не модуль. А это и означает, что данная траектория является окружностью.
Период обращения частицы в однородном магнитном поле равен:
Последнее выражение показывает, что период обращения частицы в однородном магнитном поле не зависит от скорости и радиуса траектории ее движения.
Если напряженность электрического поля равна нулю, то сила Лоренца л равна магнитной силе м :
Явление электромагнитной индукции открыл Фарадей, который установил, что в замкнутом проводящем контуре возникает электрический ток при любом изменении магнитного поля, пронизывающего контур.
Магнитный поток Ф (поток магнитной индукции) через поверхность площадью S – величина, равная произведению модуля вектора магнитной индукции на площадь S и косинус угла а между вектором и нормалью к поверхности:
Ф=BScos
В СИ единица магнитного потока 1 Вебер (Вб) – магнитный поток через поверхность площадью 1 м 2 , расположенную перпендикулярно направлению однородного магнитного поля, индукция которого равна 1 Тл:
Электромагнитная индукция-явление возникновения электрического тока в замкнутом проводящем контуре при любом изменении магнитного потока, пронизывающего контур.
Возникающий в замкнутом контуре, индукционный ток имеет такое направление, что своим магнитным полем противодействует тому изменению магнитного потока, которым он вызван (правило Ленца).
ЗАКОН ЭЛЕКТРОМАГНИТНОЙ ИНДУКЦИИ
Опыты Фарадея показали, что сила индукционного тока Ii в проводящем контуре прямо пропорциональна скорости изменения числа линий магнитной индукции , пронизывающих поверхность, ограниченную этим контуром.
Поэтому сила индукционного тока пропорциональна скорости изменения магнитного потока через поверхность, ограниченную контуром:
Известно, что если в цепи появился ток, это значит, что на свободные заряды проводника действуют сторонние силы. Работа этих сил по перемещению единичного заряда вдоль замкнутого контура называется электродвижущей силой (ЭДС). Найдем ЭДС индукции εi.
По закону Ома для замкнутой цепи
Так как R не зависит от , то
ЭДС индукции совпадает по направлению с индукционным током, а этот ток в соответствии с правилом Ленца направлен так, что созданный им магнитный поток противодействует изменению внешнего магнитного потока.
Закон электромагнитной индукции
ЭДС индукции в замкнутом контуре равна взятой с противоположным знаком скорости изменения магнитного потока, пронизывающего контур:
Опыт показывает, что магнитный поток Ф , связанный с контуром, прямо пропорционален силе тока в этом контуре:
Индуктивность контура L – коэффициент пропорциональности между проходящим по контуру током и созданным им магнитным потоком.
Индуктивность проводника зависит от его формы, размеров и свойств окружающей среды.
Самоиндукция – явление возникновения ЭДС индукции в контуре при изменении магнитного потока, вызванном изменением тока, проходящего через сам контур.
Самоиндукция – частный случай электромагнитной индукции.
Индуктивность – величина, численно равная ЭДС самоиндукции, возникающей в контуре при изменении силы тока в нем на единицу за единицу времени. В СИ за единицу индуктивности принимают индуктивность такого проводника, в котором при изменении силы тока на 1 А за 1 с возникает ЭДС самоиндукции 1 В. Эта единица называется генри (Гн):
ЭНЕРГИЯ МАГНИТНОГО ПОЛЯ
Явление самоиндукции аналогично явлению инерции. Индуктивность при изменении тока играет ту же роль, что и масса при изменении скорости тела. Аналогом скорости является сила тока.
Значит энергию магнитного поля тока можно считать величиной, подобной кинетической энергии тела :
Предположим, что после отключения катушки от источника,ток в цепи убывает со временем по линейному закону.
ЭДС самоиндукции имеет в этом случае постоянное значение:
где I – начальное значение тока, t – промежуток времени, за который сила тока убывает от I до 0.
За время t в цепи проходит электрический заряд q = Icpt . Так как Icp = (I + 0)/2 = I/2 , то q=It/2 . Поэтому работа электрического тока:
Эта работа совершается за счет энергии магнитного поля катушки. Таким образом, снова получаем:
Пример. Определите энергию магнитного поля катушки, в которой при токе 7,5 А магнитный поток равен 2,3*10 -3 Вб. Как изменится энергия поля, если сила тока уменьшиться вдвое?
Энергия магнитного поля катушки W 1 = LI 1 2 /2. По определению, индуктивность катушки L = Ф/I 1. Следовательно,
Ответ: энергия поля равна 8,6 Дж; при уменьшении тока вдвое она уменьшится в 4 раза.
Явление электромагнитной индукции
Электромагнитная индукция – явление возникновения тока в замкнутом проводящем контуре при изменении магнитного потока, пронизывающего его.
Явление электромагнитной индукции было открыто М. Фарадеем.
- На одну непроводящую основу были намотаны две катушки: витки первой катушки были расположены между витками второй. Витки одной катушки были замкнуты на гальванометр, а второй – подключены к источнику тока. При замыкании ключа и протекании тока по второй катушке в первой возникал импульс тока. При размыкании ключа также наблюдался импульс тока, но ток через гальванометр тек в противоположном направлении.
- Первая катушка была подключена к источнику тока, вторая, подключенная к гальванометру, перемещалась относительно нее. При приближении или удалении катушки фиксировался ток.
- Катушка замкнута на гальванометр, а магнит движется – вдвигается (выдвигается) – относительно катушки.
Опыты показали, что индукционный ток возникает только при изменении линий магнитной индукции. Направление тока будет различно при увеличении числа линий и при их уменьшении.
Сила индукционного тока зависит от скорости изменения магнитного потока. Может изменяться само поле, или контур может перемещаться в неоднородном магнитном поле.
Объяснения возникновения индукционного тока
Ток в цепи может существовать, когда на свободные заряды действуют сторонние силы. Работа этих сил по перемещению единичного положительного заряда вдоль замкнутого контура равна ЭДС. Значит, при изменении числа магнитных линий через поверхность, ограниченную контуром, в нем появляется ЭДС, которую называют ЭДС индукции.
Электроны в неподвижном проводнике могут приводиться в движение только электрическим полем. Это электрическое поле порождается изменяющимся во времени магнитным полем. Его называют вихревым электрическим полем. Представление о вихревом электрическом поле было введено в физику великим английским физиком Дж. Максвеллом в 1861 году.
Свойства вихревого электрического поля:
- источник – переменное магнитное поле;
- обнаруживается по действию на заряд;
- не является потенциальным;
- линии поля замкнутые.
Работа этого поля при перемещении единичного положительного заряда по замкнутому контуру равна ЭДС индукции в неподвижном проводнике.
Магнитный поток
Магнитным потоком через площадь ( S ) контура называют скалярную физическую величину, равную произведению модуля вектора магнитной индукции ( B ) , площади поверхности ( S ) , пронизываемой данным потоком, и косинуса угла ( alpha ) между направлением вектора магнитной индукции и вектора нормали (перпендикуляра к плоскости данной поверхности):
Обозначение – ( Phi ) , единица измерения в СИ – вебер (Вб).
Магнитный поток в 1 вебер создается однородным магнитным полем с индукцией 1 Тл через поверхность площадью 1 м 2 , расположенную перпендикулярно вектору магнитной индукции:
Магнитный поток можно наглядно представить как величину, пропорциональную числу магнитных линий, проходящих через данную площадь.
В зависимости от угла ( alpha ) магнитный поток может быть положительным ( ( alpha ) ( alpha ) > 90°). Если ( alpha ) = 90°, то магнитный поток равен 0.
Изменить магнитный поток можно меняя площадь контура, модуль индукции поля или расположение контура в магнитном поле (поворачивая его).
В случае неоднородного магнитного поля и неплоского контура магнитный поток находят как сумму магнитных потоков, пронизывающих площадь каждого из участков, на которые можно разбить данную поверхность.
Закон электромагнитной индукции Фарадея
Закон электромагнитной индукции (закон Фарадея):
ЭДС индукции в замкнутом контуре равна и противоположна по знаку скорости изменения магнитного потока через поверхность, ограниченную контуром:
Знак «–» в формуле позволяет учесть направление индукционного тока. Индукционный ток в замкнутом контуре имеет всегда такое направление, чтобы магнитный поток поля, созданного этим током сквозь поверхность, ограниченную контуром, уменьшал бы те изменения поля, которые вызвали появление индукционного тока.
Если контур состоит из ( N ) витков, то ЭДС индукции:
Сила индукционного тока в замкнутом проводящем контуре с сопротивлением ( R ) :
При движении проводника длиной ( l ) со скоростью ( v ) в постоянном однородном магнитном поле с индукцией ( vec ) ЭДС электромагнитной индукции равна:
где ( alpha ) – угол между векторами ( vec ) и ( vec ) .
Возникновение ЭДС индукции в движущемся в магнитном поле проводнике объясняется действием силы Лоренца на свободные заряды в движущихся проводниках. Сила Лоренца играет в этом случае роль сторонней силы.
Движущийся в магнитном поле проводник, по которому протекает индукционный ток, испытывает магнитное торможение. Полная работа силы Лоренца равна нулю.
Количество теплоты в контуре выделяется либо за счет работы внешней силы, которая поддерживает скорость проводника неизменной, либо за счет уменьшения кинетической энергии проводника.
Важно!
Изменение магнитного потока, пронизывающего замкнутый контур, может происходить по двум причинам:
- магнитный поток изменяется вследствие перемещения контура или его частей в постоянном во времени магнитном поле. Это случай, когда проводники, а вместе с ними и свободные носители заряда, движутся в магнитном поле;
- вторая причина изменения магнитного потока, пронизывающего контур, – изменение во времени магнитного поля при неподвижном контуре. В этом случае возникновение ЭДС индукции уже нельзя объяснить действием силы Лоренца. Явление электромагнитной индукции в неподвижных проводниках, возникающее при изменении окружающего магнитного поля, также описывается формулой Фарадея.
Таким образом, явления индукции в движущихся и неподвижных проводниках протекают одинаково, но физическая причина возникновения индукционного тока оказывается в этих двух случаях различной:
- в случае движущихся проводников ЭДС индукции обусловлена силой Лоренца;
- в случае неподвижных проводников ЭДС индукции является следствием действия на свободные заряды вихревого электрического поля, возникающего при изменении магнитного поля.
Правило Ленца
Направление индукционного тока определяется по правилу Ленца: индукционный ток, возбуждаемый в замкнутом контуре при изменении магнитного потока, всегда направлен так, что создаваемое им магнитное поле препятствует изменению магнитного потока, вызывающего индукционный ток.
Алгоритм решения задач с использованием правила Ленца:
- определить направление линий магнитной индукции внешнего магнитного поля;
- выяснить, как изменяется магнитный поток;
- определить направление линий магнитной индукции магнитного поля индукционного тока: если магнитный поток уменьшается, то они сонаправлены с линиями внешнего магнитного поля; если магнитный поток увеличивается, – противоположно направлению линий магнитной индукции внешнего поля;
- по правилу буравчика, зная направление линий индукции магнитного поля индукционного тока, определить направление индукционного тока.
Правило Ленца имеет глубокий физический смысл – оно выражает закон сохранения энергии.
Самоиндукция
Самоиндукция – это явление возникновения ЭДС индукции в проводнике в результате изменения тока в нем.
При изменении силы тока в катушке происходит изменение магнитного потока, создаваемого этим током. Изменение магнитного потока, пронизывающего катушку, должно вызывать появление ЭДС индукции в катушке.
В соответствии с правилом Ленца ЭДС самоиндукции препятствует нарастанию силы тока при включении и убыванию силы тока при выключении цепи.
Это приводит к тому, что при замыкании цепи, в которой есть источник тока с постоянной ЭДС, сила тока устанавливается через некоторое время.
При отключении источника ток также не прекращается мгновенно. Возникающая при этом ЭДС самоиндукции может превышать ЭДС источника.
Явление самоиндукции можно наблюдать, собрав электрическую цепь из катушки с большой индуктивностью, резистора, двух одинаковых ламп накаливания и источника тока. Резистор должен иметь такое же электрическое сопротивление, как и провод катушки.
Опыт показывает, что при замыкании цепи электрическая лампа, включенная последовательно с катушкой, загорается несколько позже, чем лампа, включенная последовательно с резистором. Нарастанию тока в цепи катушки при замыкании препятствует ЭДС самоиндукции, возникающая при возрастании магнитного потока в катушке.
При отключении источника тока вспыхивают обе лампы. В этом случае ток в цепи поддерживается ЭДС самоиндукции, возникающей при убывании магнитного потока в катушке.
ЭДС самоиндукции ( varepsilon_ ) , возникающая в катушке с индуктивностью ( L ) , по закону электромагнитной индукции равна:
ЭДС самоиндукции прямо пропорциональна индуктивности катушки и скорости изменения силы тока в катушке.
Индуктивность
Электрический ток, проходящий по проводнику, создает вокруг него магнитное поле. Магнитный поток ( Phi ) через контур из этого проводника пропорционален модулю индукции ( vec ) магнитного поля внутри контура, а индукция магнитного поля, в свою очередь, пропорциональна силе тока в проводнике.
Следовательно, магнитный поток через контур прямо пропорционален силе тока в контуре:
Индуктивность – коэффициент пропорциональности ( L ) между силой тока ( I ) в контуре и магнитным потоком ( Phi ) , создаваемым этим током:
Индуктивность зависит от размеров и формы проводника, от магнитных свойств среды, в которой находится проводник.
Единица индуктивности в СИ – генри (Гн). Индуктивность контура равна 1 генри, если при силе постоянного тока 1 ампер магнитный поток через контур равен 1 вебер:
Можно дать второе определение единицы индуктивности: элемент электрической цепи обладает индуктивностью в 1 Гн, если при равномерном изменении силы тока в цепи на 1 ампер за 1 с в нем возникает ЭДС самоиндукции 1 вольт.
Энергия магнитного поля
При отключении катушки индуктивности от источника тока лампа накаливания, включенная параллельно катушке, дает кратковременную вспышку. Ток в цепи возникает под действием ЭДС самоиндукции.
Источником энергии, выделяющейся при этом в электрической цепи, является магнитное поле катушки.
Для создания тока в контуре с индуктивностью необходимо совершить работу на преодоление ЭДС самоиндукции. Энергия магнитного поля тока вычисляется по формуле:
Основные формулы раздела «Электромагнитная индукция»
Алгоритм решения задач по теме «Электромагнитная индукция»:
1. Внимательно прочитать условие задачи. Установить причины изменения магнитного потока, пронизывающего контур.
2. Записать формулу:
- закона электромагнитной индукции;
- ЭДС индукции в движущемся проводнике, если в задаче рассматривается поступательно движущийся проводник; если в задаче рассматривается электрическая цепь, содержащая источник тока, и возникающая на одном из участков ЭДС индукции, вызванная движением проводника в магнитном поле, то сначала нужно определить величину и направление ЭДС индукции. После этого задача решается по аналогии с задачами на расчет цепи постоянного тока с несколькими источниками.
3. Записать выражение для изменения магнитного потока и подставить в формулу закона электромагнитной индукции.
4. Записать математически все дополнительные условия (чаще всего это формулы закона Ома для полной цепи, силы Ампера или силы Лоренца, формулы кинематики и динамики).
5. Решить полученную систему уравнений относительно искомой величины.
Методы созревания шейки матки и индукции родов
1. Норвиц Э., Робинсон Дж., Репке Дж. Роды и родоразрешение. В: Габби С.Г., Нибил Дж. Р., Симпсон Дж. Л., ред. Акушерство: нормальная и проблемная беременность. 4-е изд. Нью-Йорк: Черчилль Ливингстон, 2002: 353–94 ….
2. Американский колледж акушеров и гинекологов. Индукция родов. Бюллетень практики № 10. Вашингтон, округ Колумбия: ACOG, 1999.
3. Людмир Дж., Sehdev HM. Анатомия и физиология шейки матки. Clin Obstet Gynecol . 2000; 43: 433–9.
4. Эдвардс РК, Ричардс Д.С. Предварительная оценка шейки матки. Clin Obstet Gynecol . 2000; 43: 440–6.
5. МакФарлин Б.Л., Гибсон MH, О’Рир J, Харман П. Национальное исследование использования травяных препаратов медсестрами-акушерками для стимуляции родов. J Медсестра-акушерка . 1999; 44: 205–16.
6. Белью К. Травы и беременная женщина.Рекомендации для акушерок. J Медсестра-акушерка . 1999; 44: 231–52.
7. Adair CD. Нефармакологические подходы к затравке шейки матки и индукции родов. Clin Obstet Gynecol . 2000. 43: 447–54.
8. Келли А.Дж., Кавана Дж., Томас Дж. Касторовое масло, ванна и / или клизма для затравки шейки матки и индукции родов. Кокрановская база данных Syst Rev . 2002; 2: CD003099
9. Кавана Дж., Келли Эй Джей, Томас Дж.Половой акт для созревания шейки матки и индукции родов. Кокрановская база данных Syst Rev . 2002; 2: CD003093
10. Бил М.В. Иглоукалывание и точечный массаж. Приложения к охране репродуктивного здоровья женщин. J Медсестра-акушерка . 1999; 44: 217–30.
11. Smith CA, Crowther CA. Иглоукалывание для стимуляции родов. Кокрановская база данных Syst Rev . 2002; 2: CD002962
12. Lin A, Купферминц М, Дули С.Л.Рандомизированное испытание экстраамниотического солевого раствора по сравнению с ламинарией для созревания шейки матки. Акушерский гинекол . 1995. 86 (4 часть 1): 545–9.
13. Рубен Д., Ариас Ф. Рандомизированное исследование экстраамниотической инфузии физиологического раствора плюс внутрицервикальный баллон с катетером Фолея по сравнению с вагинальным гелем с простагландином E2 для созревания шейки матки и стимулирования родов у пациентов с неблагоприятными шейками матки. Акушерский гинекол . 1993; 82: 290–4.
14. Sherman DJ, Френкель Э, Панский М, Каспи Э, Буковский I, Лангер Р.Созревание шейки матки в баллоне с экстраамниотической инфузией физиологического раствора или простагландина E2: двойное слепое рандомизированное контролируемое исследование. Акушерский гинекол . 2001; 97: 375–80.
15. Buccellato CA, Стика CS, Frederiksen MC. Рандомизированное испытание мизопростола по сравнению с введением внеамниотического раствора хлорида натрия с окситоцином для индукции родов. Am J Obstet Gynecol . 2000; 182: 1039–44.
16. Гольдман Дж. Б., Wigton TR.Рандомизированное сравнение экстраамниотического солевого раствора и интрацервикального геля динопростона для созревания шейки матки. Акушерский гинекол . 1999; 93: 271–4.
17. Гуинн Д.А., Goepfert AR, Кристина М, Оуэн Дж. Hauth JC. Экстраамниотический солевой раствор, ламинария или гель простагландина E (2) для индукции родов с неблагоприятной шейкой матки: рандомизированное контролируемое исследование. Акушерский гинекол . 2000; 96: 106–12.
18. Шрейер П., Шерман DJ, Ариэли С, Герман А, Каспи Э.Созревание крайне неблагоприятной шейки матки с помощью экстраамниотического солевого раствора или вагинального применения простагландина E2. Акушерский гинекол . 1989; 73: 938–42.
19. Хади Х. Созревание шейки матки и индукция родов: клинические рекомендации. Clin Obstet Gynecol . 2000. 43: 524–36.
20. Foong LC, Ваная К., Тан G, Чуа С. Подметание мембраны в сочетании с индукцией родов. Акушерский гинекол .2000; 96: 539–42.
21. Брикер Л, Лукас М. Только амниотомия для индукции родов. Кокрановская база данных Syst Rev . 2002; 2: CD002862
22. Witter FR. Препараты простагландина Е2 для прединдукционного созревания шейки матки. Clin Obstet Gynecol . 2000; 43: 469–74.
23. Ариас Ф. Фармакология окситоцина и простагландинов. Clin Obstet Gynecol . 2000. 43: 455–68.
24. Келли А.Дж., Кавана Дж., Томас Дж.Вагинальный простагландин (PGE2 и PGF2a) для индукции родов в срок. Кокрановская база данных Syst Rev . 2002; 2: CD003101
25. Goldberg AB, Гринберг МБ, Дарни П.Д. Мизопростол и беременность. N Engl J Med . 2001; 344: 38–47.
26. Американский колледж акушеров и гинекологов. Индукция родов мизопростолом. Мнение комитета ACOG 228. Вашингтон, округ Колумбия: ACOG, 1999: 2.
27. Lydon-Rochelle M, Холт В.Л., Истерлинг TR, Мартин Д.П.Риск разрыва матки во время родов у женщин, которым ранее было выполнено кесарево сечение. N Engl J Med . 2001; 345: 3–8.
28. Sanchez-Ramos L, Годье FL, Кауниц AM. Созревание шейки матки и индукция родов после предыдущего кесарева сечения. Clin Obstet Gynecol . 2000; 43: 513–23.
29. Венгалил С.Р., Гуинн Д.А., Олаби Н.Ф., Бурд Л.И., Оуэн Дж. Рандомизированное испытание мизопростола и экстраамниотического солевого раствора для созревания шейки матки и индукции родов. Акушерский гинекол . 1998. 91 (5 часть 1): 774–9.
30. Hofmeyr GJ, Гюльмезоглу AM. Вагинальный мизопростол для созревания шейки матки и индукции родов. Кокрановская база данных Syst Rev . 2002; 2: CD000941
31. Sanchez-Ramos L, Кауниц AM. Мизопростол для созревания шейки матки и индукции родов: систематический обзор литературы. Clin Obstet Gynecol . 2000. 43: 475–88.
32. Neilson JP. Мифепристон для индукции родов. Кокрановская база данных Syst Rev . 2002; 2: CD002865
33. Келли А.Дж., Кавана Дж., Томас Дж. Релаксин для созревания шейки матки и индукции родов. Кокрановская база данных Syst Rev . 2002; 2: CD003103
34. Zeeman GG, Хан-Дауд Ф.С., Дауд М.Ю. Окситоцин и его рецепторы во время беременности и родов: современные концепции и клиническое значение. Акушерский гинекол . 1997. 89 (5 pt 2): 873–83.
35.Стаббс TM. Окситоцин для стимуляции родов .. Clin Obstet Gynecol . 2000; 43: 489–94.
Повышение квалификации и увеличение числа рабочих мест | Michigan Medicine
Обзор темы
Стимулирование родов
По мере приближения конца беременности шейка матки обычно становится мягкой (созревает), начинает открываться (расширяться) и тоньше (стираться), готовясь к схваткам и родам. Когда роды не начинаются естественным образом сами по себе и должны произойти вскоре вагинальные роды, роды могут быть начаты искусственно (индуцированно).
Несмотря на то, что стимулирование родов является довольно распространенной практикой, воспитатели по вопросам родовспоможения поощряют женщин узнавать об этом и о лекарстве для стимуляции заторможенных родов (аугментации), чтобы женщины могли помочь решить, что им подходит.
Когда роды вызваны по медицинским показаниям, обычно это происходит потому, что для вас безопаснее иметь ребенка сейчас, а не рисковать дальнейшими проблемами из-за продолжения беременности.
Ваши роды могут быть вызваны по одной из следующих причин:
- Ваша беременность прошла на 1-2 недели после предполагаемого срока родов.
- У вас есть заболевание (например, высокое кровяное давление, отслойка плаценты, инфекция, заболевание легких, преэклампсия или диабет), которое может угрожать вашему здоровью или здоровью вашего ребенка в случае продолжения беременности. сноска 1
- Вода (амниотический мешок) разорвалась, но активные схватки не начались.
- Состояние вашего ребенка требует лечения, и риск естественных родов невелик. Индукция и вагинальные роды не предпринимаются, если ребенку может быть причинен вред или он находится в непосредственной опасности.В таких случаях обычно делают кесарево сечение (кесарево сечение).
Некоторые женщины просят стимулировать роды, если для этого нет медицинских причин (индукция по желанию). А иногда врачи вызывают роды по немедицинским причинам, например, если вы живете далеко от больницы и можете не добраться до больницы, если у вас начнутся схватки. В таких ситуациях врач подождет, пока вам не исполнится 39 недель, потому что это наиболее безопасно для вашего ребенка.
Когда роды не происходят должным образом или не так, как нужно, стимулирование родов предпочтительнее родоразрешения путем кесарева сечения.Если индукция родов не увенчалась успехом, возможна еще одна попытка. В некоторых случаях кесарево сечение лучше всего для матери и ребенка, в зависимости от их состояния.
Способы стимулирования родов
Есть несколько способов вызвать родовые схватки.
- Можно использовать лекарство, чтобы смягчить шейку матки и сделать ее истонченной (стереть).
- Можно использовать лекарство, чтобы вызвать сокращение матки.
- Баллонный катетер (например, катетер Фолея) может использоваться, чтобы помочь шейке матки открыться.
- Если шейка матки мягкая и слегка приоткрытая, очистка плодных оболочек или разрыв амниотического мешка (амниотомия) может вызвать или усилить схватки.
Лекарство для созревания шейки матки и стимулирования родов
- Мизопростол (Cytotec) — это таблетка, которую принимают внутрь или вводят во влагалище (используя меньшую дозу). Это лекарство, которое в настоящее время одобрено для лечения язв. Использование этого лекарства для созревания шейки матки является широко распространенным, но немаркированным применением этого лекарства.
- Окситоцин (питоцин) можно вводить через вену (внутривенно) в небольших количествах для созревания шейки матки.Но обычно это делается после того, как шейка матки размягчается, чтобы вызвать сокращение матки. Роды, вызванные окситоцином, обычно начинаются тяжелее и прогрессируют быстрее, чем роды, которые начинаются сами по себе, особенно у рожениц впервые. Если окситоцин не вызывает родов или если частота сердечных сокращений ребенка указывает на дистресс, может потребоваться кесарево сечение (кесарево сечение).
- Динопростон (например, Cervidil или Prepidil Gel) можно вводить в виде суппозитория во влагалище (интравагинально).Его также можно вводить в виде геля, который аккуратно впрыскивают в отверстие шейки матки (интрацервикально). Когда шейка матки созрела, роды могут начаться сами по себе.
Считается, что шейка матки созрела и готова к активным родам, если она мягкая, хорошо расширенная и стертая, а также когда шейка матки и ребенок расположены низко в тазу. Если шейка матки еще не созрела, прием лекарств можно продолжать до ее созревания.
Баллонный катетер для стимулирования родов
Баллонный катетер, например катетер Фолея, представляет собой узкую трубку с небольшим баллоном на конце.Врач вставляет его в шейку матки и надувает баллон. Это помогает шейке матки открываться (расширяться). Катетер оставляют на месте до тех пор, пока шейка матки не откроется настолько, чтобы баллон выпал (около 3 см).
Очистка плодных оболочек для стимулирования родов
Очистка или удаление околоплодных вод — это простой первый шаг, используемый для попытки вызвать роды. Подметание плодных оболочек достаточно отделяет амниотическую оболочку от матки, так что матка начинает вырабатывать простагландины.Этот тип химического вещества помогает вызвать схватки и роды. После того, как шейка матки немного приоткрылась, это можно легко сделать в кабинете вашего врача или медсестры-акушерки.
Очистка плодных оболочек работает у 1 из 8 женщин. Это означает, что роды начинаются без использования окситоцина или искусственного разрыва плодных оболочек. сноска 2 Для очистки плодных оболочек врач или медсестра-акушерка протыкают шейку матки пальцем в перчатке. Затем он или она «проводит» пальцем по внутреннему краю отверстия.
Подметание мембран — низкий риск. Это не повышает риск заражения. После этого вы можете начать чувствовать дискомфорт из-за нерегулярных сокращений и небольшого кровотечения. сноска 2
Искусственный разрыв плодных оболочек для стимулирования родов
Чтобы ускорить или ускорить роды, врач может разорвать амниотический мешок (разрыв плодных оболочек). Это следует делать только после того, как шейка матки начала открываться (расширяться) и голова ребенка плотно опускается (входит) в ваш таз.При слишком раннем разрыве плодных оболочек существует риск того, что пуповина соскользнет вокруг или ниже головы ребенка (выпадение пуповины). Если пуповина зажата между головой ребенка и тазовыми костями, кровоснабжение ребенка может уменьшиться или прекратиться.
Чтобы разорвать амниотический мешок (амниотомия), врач вводит стерильное пластиковое приспособление во влагалище. Это устройство может выглядеть как длинный крючок для вязания или может быть крючком меньшего размера, прикрепленным к пальцу стерильной перчатки.Крючком осторожно натягивают амниотический мешок, пока он не разорвется. Эта процедура обычно безболезненна. Большой поток жидкости обычно следует за разрывом амниотического мешка. Матка продолжает производить околоплодные воды до рождения ребенка. Таким образом, вы можете продолжать чувствовать некоторую утечку, особенно сразу после сильного сокращения.
Аугментация
Если активные роды начались сами по себе, но схватки замедлились или полностью прекратились, необходимо предпринять шаги, чтобы помочь прогрессу родов (аугментация).Увеличение будет выполнено, когда:
- Активные роды начались, но схватки слабые, нерегулярные или полностью прекратились.
- У вас начались активные роды, но амниотический мешок не разорвался сам по себе. В этом случае врач или медсестра-акушерка могут разорвать амниотический мешок (амниотомия), чтобы ускорить роды. Если роды по-прежнему не прогрессируют, можно назначить окситоцин (питоцин), чтобы матка сократилась.
- Начались активные роды, амниотический мешок разорвался сам по себе, но роды все еще не прогрессируют.Окситоцин (питоцин) может быть введен для сокращения матки.
Если роды не развиваются, несмотря на очистку мембран, амниотомию, окситоцин или комбинацию этих мер, можно рассмотреть возможность родоразрешения путем кесарева сечения.
Методы индукции для женщин, ранее перенесших кесарево сечение
В этот обновленный обзор включены восемь исследований (данные от 707 женщин и младенцев). Мета-анализ был невозможен, поскольку в исследованиях сравнивались различные методы индукции родов.Все включенные исследования имели по крайней мере одно ограничение дизайна (например, отсутствие слепого анализа, истощение выборки, другие ошибки или систематические ошибки в отчетности). Одно исследование было преждевременно прекращено из соображений безопасности.
Вагинальный PGE2 по сравнению с внутривенным введением окситоцина (одно испытание, 42 женщины): нет четких различий для кесарева сечения (отношение рисков (ОР) 0,67, 95% доверительный интервал (ДИ) 0,22–2,03, степень доказательности низкая), серьезная неонатальная заболеваемость или перинатальный период смерть (ОР 3,00, 95% ДИ от 0,13 до 69,70, степень доказательности низкая), серьезная материнская заболеваемость или смерть (ОР 3.00, 95% ДИ от 0,13 до 69,70, степень доказательности низкая). Также нет четких различий между группами по сообщенным вторичным исходам. Результаты по шкале GRADE не были достигнуты в течение 24 часов, а гиперстимуляция матки с изменениями частоты сердечных сокращений плода не сообщалась.
Вагинальный мизопростол по сравнению с внутривенным введением окситоцина (одно испытание, 38 женщин): это испытание было прекращено досрочно, поскольку у одной женщины, получавшей мизопростол, произошел разрыв матки (ОР 3,67, 95% ДИ от 0,16 до 84,66), а у одной — расхождение матки.О других исходах (включая результаты GRADE) не сообщалось.
Катетер Фолея по сравнению с внутривенным введением окситоцина (одно испытание, подгруппа из 53 женщин): нет четкой разницы между группами по вагинальным родам, не достигнутым в течение 24 часов (ОР 1,47, 95% ДИ 0,89–2,44, степень доказательности низкая), гиперстимуляция матки с участием плода изменения частоты сердечных сокращений (ОР 3,11, 95% ДИ от 0,13 до 73,09, степень доказательности низкая) и кесарево сечение (ОР 0,93, 95% ДИ от 0,45 до 1,92, степень доказательности низкая). Также не было четких различий между группами по сообщенным вторичным исходам.Не сообщалось о следующих исходах GRADE: серьезная неонатальная заболеваемость или перинатальная смерть и серьезная материнская заболеваемость или смерть.
Двухбаллонный катетер по сравнению с вагинальным PGE2 (одно испытание, подгруппа из 26 женщин): нет четкой разницы при проведении кесарева сечения (ОР 0,97, 95% ДИ 0,41–2,32, степень доказательности очень низкая). Не сообщалось о вагинальных родах в течение 24 часов, гиперстимуляции матки с изменениями частоты сердечных сокращений плода, серьезной неонатальной заболеваемости или перинатальной смерти, а также серьезной материнской заболеваемости или смерти.
Пероральный мифепристон в сравнении с катетером Фолея (одно испытание, 107 женщин): первичных результатов / GRADE не сообщалось. Меньшему количеству женщин, индуцированных мифепристоном, потребовалось увеличение окситоцина (ОР 0,54, 95% ДИ от 0,38 до 0,76). Было немного меньше случаев разрыва матки среди женщин, получавших мифепристон, однако это не было четкой разницей между группами (ОР 0,29, 95% ДИ 0,08–1,02). О других вторичных исходах не сообщалось.
Вагинальный мононитрат изосорбида (IMN) по сравнению с катетером Фолея (одно испытание, 80 женщин): у меньшего числа женщин с индуцированным IMN роды через естественные родовые пути достигаются в течение 24 часов (RR 2.62, 95% доверительный интервал от 1,32 до 5,21, степень доказательности низкая). Не было различий между группами по количеству женщин, которым было выполнено кесарево сечение (ОР 1,00, 95% ДИ от 0,39 до 2,59, степень доказательности очень низкая). Большему количеству женщин с индуцированной IMN потребовалось увеличение окситоцина (ОР 1,65, 95% ДИ 1,17–2,32). Не было явных различий по другим зарегистрированным вторичным исходам. Не сообщалось о следующих исходах GRADE: гиперстимуляция матки с изменениями частоты сердечных сокращений плода, серьезная неонатальная заболеваемость или перинатальная смерть, а также серьезная материнская заболеваемость или смерть.
Катетер Фолея на 80 мл и 30 мл кесарево сечение (ОР 1,05, 95% ДИ 0,89–1,24, степень доказательности умеренная). Однако большему количеству женщин, индуцированных катетером Фолея на 30 мл, потребовалось увеличение окситоцина (ОР 0,81, 95% ДИ от 0,66 до 0,98). Не было четких различий между группами по другим сообщенным вторичным исходам.О некоторых исходах GRADE не сообщалось: гиперстимуляция матки с изменениями частоты сердечных сокращений плода, серьезная неонатальная заболеваемость или перинатальная смерть, а также серьезная материнская заболеваемость или смерть.
Вагинальный пессарий с PGE2 по сравнению с вагинальными таблетками с PGE2 (одно испытание, 200 женщин): нет различий между группами для кесарева сечения (ОР 1,09, 95% ДИ от 0,74 до 1,60, доказательства очень низкие) или любой из сообщенных вторичных исходов. О некоторых исходах GRADE не сообщалось: естественные роды, не достигнутые в течение 24 часов, гиперстимуляция матки с изменениями частоты сердечных сокращений плода, серьезная неонатальная заболеваемость или перинатальная смерть, а также серьезная материнская заболеваемость или смерть.
Индукция труда | GLOWM
Немедицинские методы
Были использованы многочисленные немедицинские методы для созревания шейки матки и индукции родов (Таблица 1). Хотя они популярны среди акушерок, большинство из них обычно не используются акушерами, возможно, потому, что они не были предметом должным образом проведенных рандомизированных исследований.
Таблица 1. Немедицинские методы созревания шейки матки и индукции родов
Половой акт
Стимуляция груди
Травяные препараты
Гомеопатические растворы
Слабительные средства
Клизмы
Иглоукалывание
Удаление плодных оболочек и полового акта
Существуют разумные доказательства того, что половой акт стимуляция может быть эффективной для созревания шейки матки и стимулирования родов в срок. 18 , 19 Из-за неконтролируемой секреции простагландинов и / или окситоцина, вызванной этими методами, может быть безопаснее ограничить эти подходы к доношенным женщинам со здоровой, неосложненной беременностью.
В медицинской литературе не упоминается использование растительных препаратов или гомеопатических растворов. Слабительные средства, такие как касторовое масло и клизмы, широко использовались в прошлом, но от них в основном отказались как от эффективных методов индукции родов. Иглоукалывание с ручной или электрической стимуляцией является признанным методом индукции родов в Азии и Европе; однако широко не применяется в Соединенных Штатах. 20
Удаление плодных оболочек, возможно, является наиболее изученным немедицинским методом созревания шейки матки и индукции родов. Ряд рандомизированных клинических испытаний показал, что удаление мембраны успешно вызывает роды. 21 , 22 Однако потенциальные риски включают инфекцию, преждевременный разрыв плодных оболочек и кровотечение при контакте с плацентой.
Механические методы
Механические методы, хотя в основном эффективны при раскрытии шейки матки, в течение многих лет использовались для стимулирования родов. 23 Было показано, что механическая стимуляция эндоцервикального канала запускает высвобождение простагландинов. Более популярные механические методы включают амниотомию, катетеры с баллонными наконечниками, а также натуральную и синтетическую ламинарию.
Амниотомия, или искусственный разрыв околоплодных вод, вызывает локальный синтез и высвобождение простагландинов, что приводит к родам в течение 6 часов почти у 90% доношенных пациентов. Тернбулл и Андерсон обнаружили, что амниотомия без дополнительной медикаментозной терапии успешно вызвала роды примерно в 75% случаев в течение 24 часов. 24
Механическое расширение незрелой шейки матки с помощью катетеров с баллонными наконечниками уже много лет используется для созревания шейки матки и индукции родов. Хотя описаны различные баллонные катетеры, наиболее часто используются катетеры Фолея с баллонами объемом 25–50 мл. Одновременное использование катетеров с баллонными наконечниками и фармакологических средств было эффективным при индукции родов; однако стоимость комбинированной терапии заметно возрастает. 25
Натуральная и синтетическая ламинария оказалась более эффективной для созревания шейки матки, чем индукция родов.Хотя их безопасность и эффективность были установлены во втором триместре, высокая частота инфекций связана с использованием ламинарии в третьем триместре беременности. 26
Поскольку механические агенты представляют собой инородные тела, помещенные в шейку матки или через нее, многие акушеры считают, что их использование может увеличить риск инфицирования. Недавно опубликованный метаанализ 30 рандомизированных исследований, сравнивающих индукцию родов механическими методами с альтернативными фармакологическими средствами или плацебо, показал, что материнские и неонатальные инфекции были увеличены у женщин, которым была произведена стимуляция родов механическими методами. 27 Это открытие поднимает вопрос о том, показаны ли профилактические антибиотики пациентам, подвергшимся индукции родов механическими методами.
Фармакологические методы
ОКСИТОЦИН
Окситоцин, нейрогормон, вырабатываемый в гипоталамусе и секретируемый задней долей гипофиза, представляет собой агент, наиболее часто используемый для индукции родов. Контролируемая внутривенная инфузия, с амниотомией или без нее, вызывает достаточную активность матки, чтобы вызвать расширение шейки матки и обеспечить доставку.Поскольку окситоцин часто не способствует созреванию шейки матки, он обычно не эффективен у пациентов с незрелыми шейками матки. Частота неудачных попыток индукции в этих условиях приближается к 50%, но может быть значительно снижена с использованием преиндукционных агентов для созревания шейки матки. 28
Из-за высокой активности плацентарной окситоциназы период полувыведения из плазмы короткий, и стабильные уровни достигаются после 40 минут непрерывной внутривенной инфузии. Гестационный возраст является основным фактором, влияющим на дозовую реакцию на окситоцин.Из-за появления рецепторов окситоцина в миометрии матка начинает реагировать на окситоцин примерно на 20 неделе беременности. С 34 недель беременности и до срока не отмечается изменения чувствительности. Однако, как только начинаются самопроизвольные роды, чувствительность матки быстро увеличивается.
Оптимальная начальная доза окситоцина, интервал и частота увеличения дозировки, а также методы инфузии являются предметом значительных споров. Несколько рандомизированных испытаний показали, что широкий диапазон дозировок и частот является успешным. 29 , 30 , 31 Графики увеличения дозы всего за 15 и 30 минут сравнивались с использованием начальных доз 0,5–2,5 мЕд / мин с увеличением на такое же количество; между двумя группами не было обнаружено значительных различий.
Чаще всего окситоцин начинают с дозировки 1 мЕд / мин с увеличением на 1-2 мЕд / мин каждые 20–30 минут до тех пор, пока не будет достигнута максимальная скорость введения 16-32 мЕд / мин или не будет достигнута адекватная активность матки. настоящее время.Сообщалось о других протоколах инфузии окситоцина. Более консервативный режим инфузии предусматривает начальную дозу 0,5 мЕд / мин, при этом аналогичная доза увеличивается с интервалами в 60 минут. Доказано, что 20- и 40-минутные интервалы дозирования безопасны и эффективны при использовании окситоцина в начальной дозе 6 мЕд / мин с равным увеличением.
Признание того, что эндогенный окситоцин секретируется рывками во время беременности и спонтанных родов, побудило исследовать более физиологический способ стимулирования родов с помощью этого агента.Каммиски и Дауд 32 провели рандомизированное исследование для определения безопасности и эффективности импульсного введения окситоцина по сравнению с традиционной непрерывной инфузией. Авторы пришли к выводу, что импульсное введение окситоцина так же безопасно и эффективно, как и непрерывная инфузия. Одним из очевидных преимуществ является уменьшение объема жидкости, необходимой для введения препарата, и более низкие требуемые дозы окситоцина.
Поскольку наиболее частым побочным эффектом инфузии окситоцина является замедление частоты сердечных сокращений плода (ЧСС), связанное с повышенной активностью матки, важно постоянно контролировать ЧСС и сокращения матки для выявления тахисистолии или гиперстимуляции, требующих вмешательства.Водная интоксикация, являющаяся результатом антидиуретического действия окситоцина, может возникнуть при вливании больших объемов жидкости, не содержащей электролитов.
PROSTAGLANDINS
Индукция родов с помощью простагландинов (PGs) дает преимущество в ускорении созревания шейки матки при одновременной стимуляции сократимости миометрия. Об использовании PG в качестве индукционных агентов широко сообщалось в различных классах PG, дозах и путях введения. 33 , 34 , 35 Различие между созреванием шейки матки и индукцией родов является излишним у пациентов, получающих простагландины, потому что многие женщины будут рожать после приема простагландинов.
Динопростон (PGE 2 ) — простагландин, наиболее часто используемый в акушерстве. Этот простагландин играет важную роль в процессе созревания шейки матки, а также в инициировании и поддержании родов. Оптимальный способ введения PGE 2 еще не определен. Обычно используют два пути введения: интравагинальный и интрацервикальный. Внутрицервикальный путь введения использовался примерно в двух третях клинических испытаний, о которых сообщалось. Динопростон для интрацервикального применения одобрен для коммерческого использования в США Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA) как препидил (динопростон; PGE 2 ).Коммерческий гель динопростона содержит 0,5 мг динопростона в 2,5 мл триацетина и коллоидный гель диоксида кремния в предварительно заполненном аппликаторе. Пик абсорбции препарата наступает через 30–45 минут после нанесения. Повторные дозы можно вводить с 6-часовыми интервалами, максимальная 24-часовая доза динопростона составляет 1,5 мг. Плацебо-контролируемые исследования показали, что внутрицервикальное применение PGE 2 чаще приводит к успешному созреванию шейки матки и индукции родов у пациентов с аналогичными показателями по шкале Бишопа. 36 , 37
Влагалищный вкладыш с замедленным высвобождением 10 мг динопростона также получил одобрение FDA и коммерчески доступен (Cervidil, Forest Laboratories, St Louis, MO). Вагинальный вкладыш представляет собой тонкий плоский полимерный гидрогелевый чип (29 × 9,5 × 0,8 мм) с закругленными углами, помещенный в вязаный мешок для извлечения из полиэстера. Каждая вставка содержит 10 мг динопростона в высушенной полимерной матрице, которая высвобождает динопростон с контролируемой скоростью 0.3 мг / час в течение 12 часов при регидратации при контакте со слизистой влагалища. Было показано, что вставка способствует созреванию шейки матки у беременных в срок или в ближайшем будущем, давая оценку Бишопа не менее 3 к 12 часам. Активные роды и вагинальные роды более вероятны в течение этого 12-часового периода, что снижает потребность в инфузии окситоцина. Почти три четверти пациентов нуждаются в однократном применении. 38
До утверждения FDA препаратов динопростона для интрацервикальных и вагинальных вставок часто использовался гель, приготовленный в больницах.Большинство этих препаратов сочетали в себе суппозиторий с динопростоном (Prostin E 2 , Pharmacia & Upjohn, Kalamazoo, MI) с гелем метилцеллюлозы (K-Y Jelly) и применялись либо вагинально (2,5–5 мг), либо интрацервикально (0,5 мг). Сравнительные исследования не показали каких-либо преимуществ продукта, одобренного FDA, по сравнению с гелями, приготовленными в больницах. 39 , 40
Наиболее частыми осложнениями, наблюдаемыми у пациенток, получавших PGE 2 для созревания шейки матки и индукции родов, были тахисистолия и гиперстимуляция матки.Эти результаты зависят от дозы и редко наблюдаются у пациентов, получавших небольшие дозы (0,5 мг). Другие осложнения, возникающие в результате индукции PGE 2 , включают разрыв матки, эмболию околоплодными водами и инфаркт миокарда. К счастью, эти серьезные осложнения встречаются крайне редко.
В многочисленных отчетах, включая метаанализ, было обнаружено, что мизопростол, синтетический аналог PGE 1 , безопасно и эффективно созревает шейку матки и вызывает роды у пациентов с неблагоприятными целями шейки матки. 41 Было показано, что интравагинальные дозы 25–50 мкг сокращают интервал от индукции до вагинальных родов и снижают частоту кесарева сечения. Несколько исследований показали аналогичные результаты при пероральных дозах 100 мкг каждые 4 часа. Хотя тахисистолия часто отмечается при повторных вагинальных дозах 50 мкг, частота синдрома гиперстимуляции (тахисистолия, связанная с аномалиями ЧСС) не увеличивается. Это не только безопасный и эффективный метод, но и очень экономичный.
Существуют опасения, что разделение таблеток может не обеспечить точных или согласованных доз мизопростола, и существует неопределенность в отношении характеристик высвобождения из влагалища таблетки, предназначенной для перорального применения. В настоящее время проводятся исследования по оценке безопасности и эффективности вагинального вкладыша с мизопростолом в дозах 50 и 100 мкг. В недавно опубликованном рандомизированном контролируемом исследовании сравнивали вагинальный вкладыш с мизопростолом (50 и 100 мкг) с имеющимся в продаже вагинальным вкладышем с динопростоном.Влагалищный вкладыш с мизопростолом с 100 мкг и вкладыш с динопростоном имели аналогичные средние временные интервалы с вагинальными родами. Вставка 50 мкг имела значительно более продолжительное время до вагинальных родов. 42
Другие фармакологические методы
МИФЕПРИСТОН
Роль мифепристона (RU-486), антагониста прогестерона, в индукции родов не так хорошо изучена, как для терапевтических абортов. Мифепристон с некоторым успехом использовался для индукции родов в случаях внутриутробной гибели плода на сроке не менее 16 недель.Рандомизированное двойное слепое исследование с применением 200 мг мифепристона ежедневно в течение 2 дней привело к более короткому интервалу до начала родов, и тем, у кого роды были естественным путем, требовалось меньшее количество окситоцина. 43 В группе мифепристона у 58% начались самопроизвольные роды, по сравнению с 22,6% в группе плацебо. Частота кесарева сечения не различалась между двумя группами, и никаких побочных эффектов в группе лечения не наблюдалось. Совсем недавно Elliot et al. 44 сравнили эффекты 50 мг и 200 мг мифепристона перорально с плацебо на созревание шейки матки и индукцию родов у первородящих женщин с неблагоприятными показателями шейки матки в срок.В дозе 200 мг мифепристон чаще, чем плацебо, приводил к благоприятному развитию шейки матки или самопроизвольным родам. Необходимы дальнейшие исследования, чтобы подтвердить роль мифепристона как средства, вызывающего роды.
РЕЛАКСИН
Релаксин — это полипептидный гормон, аналогичный инсулину, вырабатываемый яичниками, децидуальной оболочкой и хорионом. Поскольку он влияет на ремоделирование соединительной ткани, он был изучен как агент созревания шейки матки. Несколько клинических испытаний с использованием очищенного свиного релаксина, вводимого вагинально или интрацервикально, продемонстрировали его эффективность при созревании шейки матки.Однако недавние исследования с использованием вагинального рекомбинантного релаксина человека (1–4 мг) не показали значительного преимущества в качестве средства для преиндукционного созревания шейки матки. 45 , 46 , 47
ЦИТОКИНЫ
Роль цитокинов в созревании шейки матки в настоящее время изучается. Эти хемотаксические агенты способствуют миграции и активации воспалительных клеток, которые, в свою очередь, являются источником коллагеназы и других ферментов, способных переваривать белки внеклеточного матрикса.Было показано, что местное применение некоторых цитокинов (интерлейкин-8 [IL-8] и IL-1β) вызывает созревание шейки матки у беременных морских свинок, не вызывая явной активности матки. 48
ОКСИД АЗОТА
Исследования на животных показали, что свободный радикальный газ оксид азота активируется в шейке матки во время родов и приводит к созреванию шейки матки. 49 Недавние исследования с использованием доноров оксида азота (монотитрат изосорбида и тринитрат глицерина) показали улучшение созревания шейки матки у пациенток, прерывающих беременность в первом триместре.Роль оксида азота в созревании шейки матки и индукции родов все еще считается исследуемой. 50 В недавних публикациях оценивалась эффективность и безопасность вагинального оксида азота (изосорбид мононитрат) для амбулаторного преиндукционного созревания шейки матки. Результаты этих исследований указывают на то, что этот агент перспективен как эффективное средство для созревания шейки матки. 51 , 52
Определение неудачного индукции родов и его прогностических факторов: два нерешенных вопроса повседневной клинической ситуации — FullText — Диагностика и терапия плода 2015, Vol.38, № 3
Аннотация
Цель: Цели этого обзора заключались в том, чтобы определить прогностические факторы неудачи или успеха индукции родов (ИОЛ), а также выделить текущую неоднородность в отношении определения и диагностики неудачной ИОЛ. Материалы и методы: В обзор были включены только исследования, в которых основным или вторичным результатом была неудачная попытка ИОЛ, определяемая как не вступление в активную фазу родов после 24 часов введения простагландина ± 12 часов инфузии окситоцина.Были собраны следующие данные: дизайн исследования, определение неудачной ИОЛ, метод индукции, показания к ИОЛ, частота неудачных ИОЛ, кесарево сечение из-за неудачной ИОЛ и предикторы неудачной ИОЛ. Результатов: При поиске по базе было найдено 507 публикаций. Основная причина исключения заключалась в том, что первичные или вторичные исходы не были заранее определенным определением неудачной ИОЛ (не достижения активной фазы родов). Наконец, подходили 7 исследований. Основными прогностическими факторами, выявленными в обзоре, были статус шейки матки, оцениваемый по шкале Бишопа, или длина шейки матки. Обсуждение: Отказ ИОЛ следует определять как неспособность достичь активной фазы родов, учитывая, что определение ИОЛ — это переход в активную фазу родов. Универсальное определение неудавшейся ИОЛ является важным условием для анализа и получения надежных результатов и выводов по этому вопросу. Важным выводом этого обзора является то, что только 7 из всех рассмотренных исследований оценивали достижение активной фазы родов как первичный или вторичный исход ИОЛ. Другой вывод заключается в том, что статус шейки матки остается наиболее важным предиктором исхода ИОЛ, хотя значение параметров, исследованных до сих пор, ограничено.Поиск или разработка инструментов прогнозирования для выявления женщин, подвергшихся воздействию ИОЛ, которые могут не достичь активной фазы родов, имеет решающее значение для сведения к минимуму рисков и затрат, связанных с отказом ИОЛ, и открывает прекрасные возможности для исследования. Следовательно, необходимо изучить другие прогностические инструменты, чтобы улучшить результаты ИОЛ с точки зрения здоровья и экономического бремени.
© 2015 S. Karger AG, Базель
Введение
В последние годы индукция родов (ИОЛ) стала более частой и происходит примерно в 20% почти всех случаев [1,2].Обеспокоенность показаниями к ИОЛ, лечением и результатами растет пропорционально этому показателю. ИОЛ определяется как искусственное начало родов [3]. С другой стороны, созревание шейки матки определяется как прелюдия к началу родов, когда шейка матки становится мягкой и эластичной, что происходит либо естественным путем, либо в результате физического или фармакологического вмешательства [3]. Созревание шейки матки физическими или фармакологическими методами и индукцию родов не следует путать, хотя в литературе обычно упоминается индукция родов как процесс, включающий также созревание шейки матки.Хотя определение индукции родов простое, критерии успешной и неудачной ИОЛ не стандартизированы, и на сегодняшний день не достигнуто единого мнения. Что касается исхода ИОЛ, было предложено множество конечных точек, таких как способ родоразрешения (вагинальные роды или кесарево сечение), вагинальные роды в течение определенного временного интервала или достижение активной фазы родов. Как следствие, сравнение опубликованных исследований становится сложной задачей из-за существующей неоднородности литературы.
Определение ИОЛ, неудачной ИОЛ и кесарева сечения для неудачной ИОЛ
Некоторые авторы выразили свою озабоченность по поводу ИОЛ и определения неудачной ИОЛ [1,2]. Lin и Rouse [2] предложили практическое определение неудавшейся ИОЛ, которое должно максимизировать количество женщин, переходящих к активной фазе родов, при сохранении низкой частоты неблагоприятных исходов для матери и новорожденного. Они определяют неудачную ИОЛ как неспособность достичь раскрытия шейки матки> 4 см после 12 ± 3 часов введения окситоцина (с целью 200-225 MVU или 3 сокращений / 10 мин).Как Caughey et al. [4], упомянутые в своей статье, поскольку цель ИОЛ — вызвать у нетрудоспособной женщины начало родов, разумным определением было бы достижение активных родов в качестве меры успеха. Следовательно, это был основной результат, оцененный в нашем обзоре. Тем не менее, большинство авторов предлагают роды через естественные родовые пути в качестве основного результата ИОЛ, хотя это зависит от многих других факторов, взаимодействующих во время родов, которые не обязательно связаны с процессом индукции.
Еще один важный аспект, который следует учитывать, заключается в том, что неудачная диагностика ИОЛ в определенный момент времени не всегда подразумевает выполнение немедленного кесарева сечения во всех центрах.Это можно объяснить несколькими исследованиями, которые показали, что продолжение процесса индукции после неудачной диагностики ИОЛ в определенный момент времени в значительном количестве случаев приведет к родам через естественные родовые пути. У женщин, у которых в течение 12 часов начались активные роды, вероятность родов через естественные родовые пути составляла 67–86%, тогда как только 31–33% женщин, достигших активных родов через 18 часов, имели роды через естественные родовые пути [5]. Следовательно, неудавшаяся диагностика ИОЛ не всегда включает выполнение кесарева сечения при неудачной ИОЛ, и, следовательно, сообщаемые показатели этих двух событий различаются между исследованиями.Кроме того, в большинстве руководств не указывается ограничение по времени от инициации ИОЛ до доставки [3], а также не достигнут консенсус относительно продолжительности латентной и активной фазы. Скрытая фаза определяется как период времени, не обязательно непрерывный, когда наблюдаются болезненные сокращения и некоторые изменения шейки матки, включая сглаживание и расширение шейки матки до 4 см, и начало активных родов, когда есть регулярные болезненные схватки и прогрессирующие раскрытие шейки матки от 4 см [6].
Несмотря на отсутствие конкретных рекомендаций, определение неудачной ИОЛ, совпадающее по времени с низкой вероятностью достижения активной фазы родов, кажется лучшим вариантом. Неблагоприятные исходы, связанные с продолжительностью латентной фазы, широко не оценивались, хотя некоторые исследования связывают длительную латентную фазу с последующими аномалиями родов и необходимостью кесарева сечения [7]. Длительная латентная фаза более 12 часов также была связана со значительно большей продолжительностью активных родов [8].Тем не менее, более высокая частота хориоамнионита (от 20-22 до 25-27%) и послеродового кровотечения (от 11 до 16%) была зарегистрирована после 6 и 12 часов латентной фазы, соответственно [5,9]. Кроме того, необходимо учитывать экономические затраты, связанные со сроком действия ИОЛ [10]. Это важная информация, которая в сочетании с другими прогностическими факторами может помочь в принятии решения о целесообразности продолжения индукционного процесса.
Таким образом, важно диагностировать отказавшую ИОЛ в подходящее время, чтобы проконсультировать пациентов и решить, продолжать ли ИОЛ или выполнить кесарево сечение, исходя из низкой вероятности перехода в активную фазу родов без увеличение неблагоприятных исходов [11].Основные цели этого обзора состояли в том, чтобы определить прогностические факторы неудачи или успеха ИОЛ, а также подчеркнуть существующую неоднородность в отношении определения и диагностики неудачной ИОЛ, которая может ухудшить идентификацию прогностических факторов.
Материалы и методы
Идентификация литературы
Был проведен поиск в электронной базе данных (PubMed, MEDLINE и Embase) до января 2014 года. Стратегия поиска для идентификации исследований исходов ИОЛ и ее прогностических факторов состояла из следующей комбинации терминов MeSH: «индуцированный, родовой», «результат лечения», «ультразвуковое исследование» и «шейка матки».
Мы проверили справочные списки релевантных исследований, чтобы выявить цитируемые статьи, не обнаруженные с помощью электронного поиска, а функция «похожие статьи» в PubMed была использована для завершения поиска. Два исследователя (N.B., F.M.) независимо проверяли заголовки и выдержки из результатов электронного поиска на предмет релевантности. Полные тексты потенциально подходящих цитат были получены для дальнейшей оценки. Отбор на включение и извлечение данных проводился одним рецензентом (N.B.) и проверялся вторым рецензентом (F.М.).
Мы включили ретроспективные и проспективные исследования, а также систематические обзоры и метаанализы. Ограничений относительно показания ИОЛ или метода индукции не применялось. Исследования включались только в том случае, если их основным или вторичным результатом была неудачная попытка ИОЛ, определяемая как выход из активной фазы родов после 24 часов введения простагландина ± 12 часов инфузии окситоцина. Были собраны следующие данные: дизайн исследования, точное определение неудачной ИОЛ, метод индукции, показание ИОЛ, частота неудачных ИОЛ, кесарево сечение из-за неудачной ИОЛ и предикторы неудачной ИОЛ.
Результаты
Описание исследований
При поиске по базе было обнаружено 507 публикаций. После первого обзора названий и аннотаций 427 были исключены. Получены полные тексты 80 статей. Основная причина исключения заключалась в том, что первичные или вторичные исходы не были общепринятым определением неудачной ИОЛ (недостижение активной фазы родов), как показано на рисунке 1. Наконец, подходили 7 исследований. В 5 из 7 включенных исследований основным результатом оценки было достижение активной фазы родов.В двух исследованиях второстепенным исходом было прекращение активной фазы родов [12,13], и они также были включены в обзор. Характеристики включенных исследований приведены в таблице 1.
Таблица 1
Характеристики исследований, включенных в обзор
Рис. 1
Выбор исследований, включенных в обзор.
Показания для ИОЛ
Медицинские и плановые ИОЛ следует различать, поскольку результаты ИОЛ различаются в зависимости от показаний к индукции.При наличии медицинских показаний потенциальная польза для матери или плода легко превосходит риски, связанные с ИОЛ. Рекомендации по применению ИОЛ для переношенной беременности, преждевременного разрыва плодных оболочек в срок и преждевременного разрыва плодных оболочек в ближайшем будущем с легочной зрелостью подтверждаются доказательствами. В этих ситуациях с ИОЛ не наблюдается повышенной частоты кесарева сечения [14]. Плоды с задержкой внутриутробного развития демонстрируют более высокую внутриутробную смертность в группе беременных, уравновешенную увеличением неонатальной смертности в группе немедленной индукции.Имеется мало данных относительно инсулино-требующего диабета, беременности двойней, макросомии плода, маловодия, холестаза при беременности, сердечных заболеваний матери и гастрошизиса плода [1,14]. Переношенная беременность является наиболее частым показанием для ИОЛ. С другой стороны, выборная ИОЛ становится все более частым показанием к ИОЛ, особенно на основании последних научных данных [15]. Несмотря на низкий риск осложнений для матери и плода, связанных с плановой ИОЛ, благоприятные цервикальные и акушерские условия должны быть предварительным условием при указании плановой ИОЛ, чтобы еще больше снизить возможные неблагоприятные исходы, связанные с этим вмешательством.Показания к ИОЛ показаны в таблице 2. В исследованиях, включенных в наш обзор, частота плановой индукции варьируется от 0,95 до 10%, показывая прогрессивное увеличение почти во всех условиях. В одном исследовании [12] показаний к ИОЛ не сообщалось.
Таблица 2
Частота кесарева сечения при несостоятельности ИОЛ
Более высокий уровень материнской и плодовой заболеваемости, особенно с повышенным риском кесарева сечения, был связан с ИОЛ [16,17], но эта связь была оспорена в последнее время. годы.В Кокрановском обзоре сделан вывод о том, что частота кесарева сечения и частота искусственных родов не увеличивается при использовании ИОЛ в срок при наличии медицинских показаний [18,19]. Было продемонстрировано, что частота кесарева сечения не увеличивается или даже снижается при применении ИОЛ на 41 неделе беременности или позже, в дополнение к улучшению перинатальных исходов [15,20]. О противоречивых результатах сообщают на сроках гестации менее 40–41 недель [21,22].
Все приведенные данные говорят о том, что в ближайшие годы количество ИОЛ может еще больше увеличиться.Следовательно, чрезвычайно важно достичь консенсуса в отношении лечения этого чрезвычайно частого вмешательства.
Как упоминалось выше, диагностика неудачной ИОЛ не всегда подразумевает выполнение кесарева сечения в это время. Кроме того, не во всех исследованиях, включенных в обзор, сообщалось как о неудачной диагностике ИОЛ, так и о кесаревом сечении при неудачной оценке ИОЛ, как показано в таблице 3. Тем не менее, несмотря на ограниченное количество исследований, незначительная часть пациенток родилась через естественные родовые пути после постановки диагноза. с неудавшейся ИОЛ в 3 из 7 включенных исследований [23,24,25].
Таблица 3
Диагностика неудачной ИОЛ и кесарево сечение при неудачной ИОЛ
Факторы, определяющие исход ИОЛ
Несмотря на большое количество проведенных исследований, не было обнаружено никаких хороших прогностических факторов для успешного определения группы пациентов, которые не будут реагировать на физическая или фармакологическая индукция и, следовательно, достижение активной фазы родов. Неоднородность оцениваемых результатов способствует отсутствию убедительных результатов.
Цервикальный статус, в основном измеряемый по шкале Бишопа и паритету, оказался основным предиктором успешного индукции родов [26,27].Сообщалось о других прогностических факторах, таких как возраст матери, вес, рост, индекс массы тела, этническая принадлежность и социально-экономический статус. Также описан акушерский и медицинский анамнез, такой как гестационный возраст при родах, масса тела при рождении и индекс околоплодных вод [28]. Соответствующие прогностические факторы, представленные в исследованиях, включенных в обзор, показаны в таблице 4.
Таблица 4
Прогностические факторы исхода ИОЛ
Гестационный возраст
Плацентарная функция снижается, в то время как непереносимость родов и масса плода увеличивается с возрастом гестации. .Все эти факторы могут способствовать увеличению частоты кесарева сечения. Однако отсутствие перехода в активную фазу родов и, как следствие, неудачная индукция не связаны с тремя факторами, описанными выше [29]. В одном из включенных исследований было обнаружено, что более ранний гестационный возраст является важным прогностическим фактором неудачной ИОЛ [24].
Характеристики матери
Возраст матери способствует более высокой распространенности материнских осложнений во время беременности и, следовательно, увеличению частоты ИОЛ, в том числе при неблагоприятных условиях.Дисфункциональный миометрий также может привести к более длительным родам и, как следствие, к большему количеству неудачных ИОЛ [30]. Однако в рассмотренных исследованиях не было обнаружено значимой связи между возрастом матери и исходом ИОЛ. Более высокий индекс массы тела связан с более высоким весом плода и осложнениями, связанными с беременностью, причем оба состояния связаны с повышенным риском кесарева сечения. Однако относительно достижения самой активной фазы родов эта связь не была продемонстрирована [31,32,33].
Состояние шейки матки
Поскольку состояние шейки матки перед родами было признано наиболее важным предиктором успеха индукции, во многих исследованиях оценивалась взаимосвязь между оценкой Бишопа и сонографическими характеристиками шейки матки с исходом ИОЛ. Что касается прогностической ценности оценки Бишопа, метаанализ [34] и недавно опубликованный систематический обзор [35] дали разные результаты. Teixeira et al. [34] пришли к выводу, что оценка по шкале Бишопа по-прежнему является наиболее точным прогностическим фактором вагинальных родов после ИОЛ.Когда речь идет о достижении активной фазы родов , либо без ограничения по времени для этого, либо в течение определенного временного интервала, такой же вывод нельзя сделать из-за отсутствия исследований, оценивающих этот результат. С другой стороны, Kolkman et al. [35] показали, что оценка по Бишопу 4, 5 или 6 была плохим предиктором успеха ИОЛ, если это определялось как достижение естественных родов. Для прогноза кесарева сечения чувствительность и специфичность по шкале Бишопа <6 составила 78 и 44% соответственно, а по шкале Бишопа <9 - 95 и 30%.В этом исследовании рекомендуется не использовать шкалу Бишопа при принятии решений. В трех исследованиях, включенных в наш обзор [25,36], [37], было обнаружено, что оценка Бишопа является важным прогностическим фактором исхода ИОЛ, хотя ни в одном из исследований она не была сильным прогностическим фактором. В одном исследовании расширение шейки матки и сглаживание шейки матки были независимо связаны с «достижением активной фазы родов» [12]. Оценка Бишопа не была независимым прогностическим фактором в других включенных исследованиях (таблица 4).
Что касается сонографической оценки шейки матки для прогнозирования исхода ИОЛ, также сообщается о противоречивых результатах.Недавний метаанализ выявил ограниченное значение длины шейки матки и клиновидности шейки матки для прогнозирования исхода родов в клинической практике [38]. Более того, из 31 исследования, включенного в метаанализ, только в 2 сообщается о результате «недостижение активной фазы родов». Чувствительность и специфичность длины шейки матки в исследованиях, включенных в этот обзор, составляли 56–66% [37] и 83–75% [23], соответственно, для отсечки 30 мм. В другом систематическом обзоре Hatfield et al. [26] также пришли к выводу, что сонографическая длина шейки матки не была эффективным или, в лучшем случае, слабым предиктором любого из оцениваемых исходов (способ родоразрешения, вагинальные роды в течение 24 часов и достижение активной фазы родов).Более того, при сравнении оценки Бишопа с длиной шейки матки, измеренной с помощью УЗИ, сонографическая длина шейки матки не превосходила оценку Бишопа. Длина шейки матки оказалась независимым прогностическим фактором в 2 исследованиях, включенных в наш обзор [23,24]. Другие изученные сонографические характеристики, такие как заклинивание шейки матки (чувствительность 37%, специфичность 80%) [38] и положение головы плода (чувствительность 39%, специфичность 71%), не оказались хорошими предикторами [39].
Подводя итог, можно сказать, что существует связь между оценкой Бишопа и длиной шейки матки, измеренной с помощью УЗИ, и исходом ИОЛ, но эта связь не всегда дает хороший прогноз.Поэтому в будущих исследованиях следует изучить другие характеристики шейки матки, комбинацию факторов или сосредоточиться на новых диагностических инструментах.
Некоторые модели прогнозирования были разработаны для прогнозирования риска кесарева сечения после ИОЛ [40,41,42], хотя ни одна из этих моделей не оценивала прогноз достижения активной фазы родов. Более того, их использование в клинической практике не рекомендуется из-за их умеренной прогностической способности [43].
Другие факторы прогноза
Биохимические маркеры
Биохимические маркеры, такие как фибронектин плода и IGFBP-1, также были изучены, но ни один из них не превосходит оценку Бишопа в отношении прогноза успешной ИОЛ [44,45, 46].
Новые прогностические факторы на основе анализа изображений
Разрабатывается широкий спектр технологий и устройств для оценки характеристик шейки матки, но только два из них были оценены у женщин, перенесших ИОЛ: светоиндуцированная флуоресценция (LIF) и эластография.
Разработан метод измерения цервикального коллагена с помощью LIF [47, 48]. Fittkow et al. [49] обнаружили, что измерения LIF отрицательно коррелируют со сроком беременности и положительно со временем до родов, хотя корреляция была слабой.Они применили этот метод для оценки созревания шейки матки в условиях ИОЛ. LIF и показатель Бишопа измеряли в начале ИОЛ. Корреляции между двумя параметрами не обнаружено. Те же измерения были выполнены через 4 часа после введения простагландина, обнаружив статистически значимую обратную корреляцию. С другой стороны, эластография основана на определении движения в областях шейки матки относительно других, определяя индекс эластографии (EI). В исследовании 29 женщин, перенесших ИОЛ, у женщин с успешной ИОЛ был более высокий EI внутреннего зева, чем у женщин с неудавшейся ИОЛ.EI не был прогностическим в других цервикальных областях. Muscatello et al. [50] также сообщили о более высокой частоте кесарева сечения у женщин с более высоким EI. Однако недавнее исследование показало, что эластографическая оценка внутреннего зева вряд ли может быть полезна для прогнозирования вагинальных родов и интервала от индукции до родов [51]. Техническое ограничение заключается в том, что это измерение все еще зависит от силы, прилагаемой оператором, и, следовательно, необходимы дальнейшие исследования [52].
Разнообразные другие методы и устройства исследуются для определения состава и поведения шейки матки, хотя до сих пор они ориентированы только на прогнозирование преждевременных родов [53].Некоторыми примерами этих методов являются акустическое затухание [54] и электрическое сопротивление [55], которые оценивают гидратацию шейки матки. Генерация второй гармоники [56], спектроскопия комбинационного рассеяния [57,58] и потеря мощности обратного рассеяния [58] также изучают микроструктуру цервикального коллагена. Что касается оценки эластичности тканей, также были оценены несколько стратегий, таких как индекс консистенции шейки матки [59] и устройство для аспирации шейки матки [60]. Отсутствие области шейных желез также было предложено в качестве предиктора преждевременных родов [61,62].
Подводя итог, можно сказать, что существует большое количество исследований, описывающих новые параметры для более точной оценки изменений шейки матки во время беременности, тем самым отражая большой интерес научного сообщества к поиску объективного и применимого метода оценки для прогнозирования соответствующих клинических исходов, таких как несостоятельность ИОЛ или успех и риск преждевременных родов. К сожалению, несмотря на очень частые ситуации, надежный предсказатель пока не найден.
Обсуждение
ИОЛ — чрезвычайно распространенная процедура, выполняемая во всех акушерских учреждениях.Примерно в 20% беременностей роды вызываются по разным причинам, наиболее частым показанием является перенесенная беременность. Несмотря на огромное медицинское и экономическое воздействие, до сих пор существует много споров относительно определения ИОЛ, показаний, лечения и результатов. Неоднородность протоколов и определений ИОЛ очень затрудняет делать выводы на основе опубликованных исследований.
Первый аспект, на который следует обратить внимание, — это отсутствие общепринятого определения неудачной ИОЛ.Большинство исследований определяют неудавшуюся ИОЛ как невозможность достижения вагинальных родов, хотя во время родов появляется много других факторов, которые могут препятствовать или предотвращать вагинальные роды, что может объяснить, почему не был найден надежный предиктор. С нашей точки зрения, определение неудавшейся ИОЛ должно соответствовать самому определению ИОЛ, которое является достижением активной фазы родов. Следовательно, неудавшаяся интраокулярная линза должна определяться как невозможность достижения этого. Важным выводом этого обзора является то, что только 7 из всех рассмотренных исследований оценивали достижение активной фазы родов как первичный или вторичный исход ИОЛ.Это означает, что большая часть существующей литературы основана на очень общем результате, оценивая конечный результат, такой как вагинальные роды, и, следовательно, добавляя смешивающие факторы.
Второй важный вывод этого обзора состоит в том, что сами клинические факторы риска имеют слишком низкую прогностическую ценность, чтобы их можно было преобразовать в полезный клинический инструмент. Напротив, статус шейки матки остается наиболее важным предиктором исхода ИОЛ, хотя значение параметров, исследованных на сегодняшний день, ограничено.Следовательно, прилагаются большие усилия, чтобы найти объективный метод для более точной оценки состояния шейки матки. Поиск или разработка инструментов прогнозирования для выявления женщин, подвергшихся воздействию ИОЛ, которые могут не достичь активной фазы родов, имеет решающее значение для минимизации рисков и затрат, связанных с отказом ИОЛ.
В заключение, общепринятое и адекватное определение неудавшейся ИОЛ является важным условием для анализа и получения надежных результатов и выводов. С другой стороны, улучшение прогнозирования отказа ИОЛ в настоящее время является серьезной проблемой в акушерстве из-за негативного воздействия неудачной ИОЛ в нескольких областях.Наша неспособность предсказать это приводит к длительным и дорогостоящим индукционным трудам. Выявление таких женщин позволит получить индивидуальное консультирование и потенциально улучшить исходы с точки зрения здоровья (неблагоприятные перинатальные и материнские исходы из-за длительного индукции родов), экономического бремени (затраты на персонал, лекарства и госпитализацию) и восприятия качества системы здравоохранения (материнские дискомфорт). Следовательно, срочно необходимы новые предикторы для этого частого вмешательства, чтобы улучшить ведение ИОЛ и результаты.
Список литературы
- Талауликар В.С., Арулкумаран С: Неудачное стимулирование родов: стратегии повышения успешности. Акушерский гинекологический обзор 2011; 66: 717-728.
- Лин М.Г., Роуз DJ: Что такое неудавшаяся индукция родов? Clin Obstet Gynecol 2006; 49: 585-593.
- Бюллетень практики ACOG № 107: Индукция родов. Obstet Gynecol 2009; 114: 386-397.
- Caughey AB, Sundaram V, Kaimal AJ, Gienger A, Cheng YW, McDonald KM, Shaffer BL, Owens DK, Bravata DM: систематический обзор: плановая индукция родов по сравнению с выжидательной тактикой беременности.Энн Интерн Мед 2009; 151: 252-263, W53-63.
- Саймон CE, Гробман WA: Когда индукция не работает? Obstet Gynecol 2005; 105: 705-709.
- Королевский колледж акушерок: научно обоснованные рекомендации по оказанию акушерской помощи в родах.Скрытая фаза. Лондон, Королевский колледж акушерок, 2012 г.
- Chelmow D, Kilpatrick SJ, Laros RK Jr: Материнские и неонатальные исходы после длительной латентной фазы. Obstet Gynecol 1993; 81: 486-491.
- Dencker A, Berg M, Bergqvist L, Lilja H: Определение факторов латентной фазы, связанных с продолжительностью активных родов у первородящих женщин с низким риском спонтанных схваток.Acta Obstet Gynecol Scand 2010; 89: 1034-1039.
- Rouse DJ, Owen J, Hauth JC: Критерии неудачной индукции родов: проспективная оценка стандартизированного протокола. Obstet Gynecol 2000; 96: 671-677.
- Rouse DJ, Weiner SJ, Bloom SL, Varner MW, Spong CY, Ramin SM, Caritis SN, Grobman WA, Sorokin Y, Sciscione A, Carpenter MW, Mercer BM, Thorp JM Jr, Malone FD, Harper M, Iams JD, Anderson GD: Неудачное стимулирование родов: к объективному диагнозу.Акушерский гинекол 2011; 117: 267-272.
- Cromi A, Ghezzi F, Tomera S, Scandroglio S, Colombo G, Bolis P: Созревание шейки матки с катетером Фолея: роль ультразвукового исследования шейки матки до и после созревания. Am J Obstet Gynecol 2007; 196: 41.e1-7.
- Chandra S, Crane JM, Hutchens D, Young DC: Трансвагинальное ультразвуковое исследование и цифровое обследование в прогнозировании успешной индукции родов.Obstet Gynecol 2001; 98: 2-6.
- Фредерикс Ф., Ли С., Деккер Г. Факторы риска неудачной индукции у нерожавших женщин. J Matern Fetal Neonatal Med 2012; 25: 2479-2487.
- Mozurkewich E, Chilimigras J, Koepke E, Keeton K, King VJ: Показания для индукции родов: обзор с наилучшими доказательствами.BJOG 2009; 116: 626-636.
- Дарни Б.Г., Сноуден Дж. М., Ченг Ю. В., Джейкоб Л., Николсон Дж. М., Каймал А., Дублин С., Гетахун Д., Кауги А. Б.: Плановая индукция родов в срок по сравнению с выжидательной тактикой: исходы для матери и новорожденного. Obstet Gynecol 2013; 122: 761-769.
- Battista L, Chung JH, Lagrew DC, Wing DA: Осложнения индукции родов у повторнородящих женщин в больничной системе по месту жительства.Am J Obstet Gynecol 2007; 197: 241.e1-7, обсуждение 322-323.e1-4.
- Гривелл Р.М., Рейли А.Дж., Оки Х., Чан А., Додд Дж. М.: Материнские и неонатальные исходы после индукции родов: когортное исследование. Acta Obstet Gynecol Scand 2011; 91: 198-203.
- Гульмезоглу AM, Crowther CA, Миддлтон П: Индукция родов для улучшения исходов родов у женщин в срок или после родов.Кокрановская база данных Syst Rev 2006; CD004945.
- Dare MR, Middleton P, Crowther CA, Flenady VJ, Varatharaju B: Планируемые ранние роды по сравнению с выжидательной тактикой (ожиданием) при дородовом разрыве плодных оболочек в срок (37 недель и более). Кокрановская база данных Syst Rev 2006; CD005302.
- Сток С.Дж., Фергюсон Э., Даффи А., Форд И., Чалмерс Дж., Норман Дж. Э.: Результаты выборного индукции родов по сравнению с выжидательной тактикой: популяционное исследование.BMJ 2012; 344: e2838.
- Санчес-Рамос Л., Оливье Ф., Делке И., Кауниц А.М.: индукция родов по сравнению с выжидательной тактикой при послеродовой беременности: систематический обзор с метаанализом. Акушерский гинекол 2003; 101: 1312-1318.
- Nicholson JM, Caughey AB, Stenson MH, Cronholm P, Kellar L, Bennett I., Margo K, Stratton J: Активное управление риском многоплодной беременности в срок: связь между более высоким показателем превентивной индукции родов и улучшенными исходами родов.Am J Obstet Gynecol 2009; 200: 250.e1-250.e13.
- Ян Ш., Ро Ч. Р., Ким Дж. Х .: Трансвагинальное ультразвуковое исследование для оценки шейки матки перед индукцией родов. J Ultrasound Med 2004; 23: 375-382, викторина 384-385.
- Park KH: Трансвагинальное ультразвуковое исследование шейки матки для прогнозирования неудачной индукции родов и кесарева сечения при отсутствии прогресса у первородящих женщин.J Korean Med Sci 2007; 22: 722-727.
- Park KH, Hong JS, Shin DM, Kang WS: Прогнозирование неудачной индукции родов у рожавших женщин в срок: роль предыдущего акушерского анамнеза, цифровое обследование и ультразвуковое измерение длины шейки матки. Журнал J Obstet Gynaecol Res 2009; 35: 301-306.
- Hatfield AS, Sanchez-Ramos L, Kaunitz AM: Сонографическая оценка шейки матки для прогнозирования успеха индукции родов: систематический обзор с метаанализом. Am J Obstet Gynecol 2007; 197: 186-192.
- Гробман В.А.: Предикторы успеха индукции.Семин Перинатол 2012; 36: 344-347.
- Крейн JM: Факторы, предсказывающие успех индукции родов: критический анализ. Clin Obstet Gynecol 2006; 49: 573-584.
- Caughey AB, Stotland NE, Washington AE, Escobar GJ: Материнские и акушерские осложнения беременности связаны с увеличением срока беременности в срок.Am J Obstet Gynecol 2007; 196: 155.e1-6.
- Гринберг М.Б., Ченг Ю.В., Салливан М., Нортон М.Е., Хопкинс Л.М., Кауги А.Б.: Зависит ли продолжительность родов от возраста матери? Am J Obstet Gynecol 2007; 197: 428.e1-7.
- Nuthalapaty FS, Rouse DJ, Owen J: Связь веса матери с риском кесарева сечения, продолжительностью родов и скоростью раскрытия шейки матки во время индукции родов.Obstet Gynecol 2004; 103: 452-456.
- Ecker JL, Chen KT, Cohen AP, Riley LE, Lieberman ES: Повышенный риск кесарева сечения с увеличением возраста матери: показания и связанные факторы у нерожавших женщин. Am J Obstet Gynecol 2001; 185: 883-887.
- Weiss JL, Malone FD, Emig D, Ball RH, Nyberg DA, Comstock CH, Saade G, Eddleman K, Carter SM, Craigo SD, Carr SR, D’Alton ME: ожирение, акушерские осложнения и частота кесарева сечения — популяция- основанное на скрининговом исследовании.Ам Дж. Обстет Гинекол 2004; 190: 1091-1097.
- Teixeira C, Lunet N, Rodrigues T., Barros H: Оценка Бишопа как определяющий фактор успеха индукции родов: систематический обзор и метаанализ. Arch Gynecol Obstet 2012; 286: 739-753.
- Kolkman DG, Verhoeven CJ, Brinkhorst SJ, van der Post JA, Pajkrt E, Opmeer BC, Mol BW: Оценка Бишопа как предиктор успеха индукции родов: систематический обзор.Am J Perinatol 2013; 30: 625-630.
- Xenakis EM, Piper JM, Conway DL, Langer O: Индукция родов в девяностые: победа над неблагоприятной шейкой матки. Obstet Gynecol 1997; 90: 235-239.
- Роман Х, Верспик Э., Веркустр Л., Дегре С., Кол JY, Фирмин Дж. М., Карон П., Марпо Л.: Улучшает ли ультразвуковое исследование при неблагоприятном состоянии шейки матки прогноз неудачной индукции родов? Ультразвуковой акушерский гинекол 2004; 23: 357-362.
- Verhoeven CJ, Opmeer BC, Oei SG, Latour V, van der Post JA, Mol BW: Трансвагинальная сонографическая оценка длины шейки матки и расклинивания для прогнозирования исхода индукции родов в срок: систематический обзор и метаанализ. Ультразвуковой акушерский гинеколь 2013; 42: 500-508.
- Верхувен С.Дж., Малдерс Л.Г., Оей С.Г., Мол Б.В.: Предсказывает ли положение головы плода при ультразвуковом исследовании до индукции родов исход родов? Eur J Obstet Gynecol Reprod Biol 2012; 164: 133-137.
- Peregrine E, O’Brien P, Omar R, Jauniaux E: Клинические и ультразвуковые параметры для прогнозирования риска кесарева сечения после индукции родов. Obstet Gynecol 2006; 107: 227-233.
- Рэйн С.М., Гирджис Р.Р., Хиггинс Б., Николаидес К.Х.: Модели для прогнозирования успешной индукции родов, основанные на предварительном сонографическом измерении длины шейки матки.J Matern Fetal Neonatal Med 2005; 17: 315-322.
- Кипанассерил А., Сури В., Багга Р., Аггарвал Н.: Новая система объективной оценки для прогнозирования успешного индукции родов. Журнал J Obstet Gynaecol 2012; 32: 145-147.
- Verhoeven CJ, Oudenaarden A, Hermus MA, Porath MM, Oei SG, Mol BW: Валидация моделей, прогнозирующих кесарево сечение после индукции родов.Ультразвуковой акушерский гинеколь 2009; 34: 316-321.
- Reis FM, Gervasi MT, Florio P, Bracalente G, Fadalti M, Severi FM, Petraglia F: Прогнозирование успешного индукции родов в срок: роль анамнеза, цифровое обследование, ультразвуковая оценка шейки матки и анализ фибронектина плода.Am J Obstet Gynecol 2003; 189: 1361-1367.
- Ojutiku D, Jones G, Bewley S: Количественный фибронектин плода как предиктор успешного индукции родов при послеродовой беременности. Eur J Obstet Gynecol Reprod Biol 2002; 101: 143-146.
- Nuutila M, Hiilesmaa V, Karkkainen T, Ylikorkala O, Rutanen EM: Фосфорилированные изоформы белка-1, связывающего инсулиноподобный фактор роста, в шейке матки как предиктор созревания шейки матки.Obstet Gynecol 1999; 94: 243-249.
- Мол Х, Сааде Дж., Гарфилд РЭ: Прогнозирование сроков и преждевременных родов и мониторинг лечения путем измерения сшитого коллагена шейки матки с использованием индуцированной светом флуоресценции. Acta Obstet Gynecol Scand 2005; 84: 534-536.
- Schlembach D, Mackay L, Shi L, Maner WL, Garfield RE, Maul H: Созревание шейки матки и недостаточность: от биохимических и молекулярных исследований до клинического обследования in vivo.Eur J Obstet Gynecol Reprod Biol 2009; 144 (приложение 1): S70-S76.
- Фитткоу CT, Maul H, Olson G, Martin E, MacKay LB, Saade GR, Garfield RE: светоиндуцированная флуоресценция шейки матки человека уменьшается после применения простагландина для индукции родов в срок. Eur J Obstet Gynecol Reprod Biol 2005; 123: 62-66.
- Muscatello A, Di Nicola M, Accurti V, Mastrocola N, Franchi V, Colagrande I, Patacchiola F, Carta G: соноэластография как метод предварительной оценки шейки матки для прогнозирования успеха индукции родов. Fetal Diagn Ther 2014; 35: 57-61.
- Pereira S, Frick AP, Poon LC, Zamprakou A, Nicolaides KH: Успешное индукция родов: прогнозирование по длине шейки матки до индукции, углу прогрессирования и эластографии шейки матки.Ультразвуковой акушерский гинеколь 2014; 44: 468-475.
- Святковска-Фройнд М., Прейс К.: Эластография шейки матки: значение для успеха индукции родов. Ультразвуковой акушерский гинеколь 2011; 38: 52-56.
- Feltovich H, Hall TJ, Berghella V: За пределами длины шейки матки: новые технологии для оценки шейки матки беременных.Am J Obstet Gynecol 2012; 207: 345-354.
- McFarlin BL, Bigelow TA, Laybed Y, O’Brien WD, Oelze ML, Abramowicz JS: Оценка ослабления ультразвука шейки матки беременной: предварительный отчет. Ультразвуковой акушерский гинеколь 2010; 36: 218-225.
- Джохи Р.П., Браун Б.Х., Анумба Д.О .: Роль спектроскопии электрического импеданса шейки матки в прогнозировании течения и исхода искусственных родов.BMC Беременность и роды 2009; 9:40.
- Акинс М.Л., Луби-Фелпс К., Махендру М.: Визуализация генерации второй гармоники как потенциальный инструмент для определения стадии беременности и прогнозирования преждевременных родов. Дж. Биомед Опт 2010; 15: 026020.
- Варгис Э., Берд Т., Логан К., Хабеле Д., Махадеван-Янсен А.: Чувствительность рамановской спектроскопии к нормальной вариабельности пациента.Дж. Биомед Опт 2011; 16: 117004.
- Фельтович Х., Нам К., Холл Т.Дж .: Количественная ультразвуковая оценка микроструктуры шейки матки. Ультразвуковая визуализация 2010; 32: 131-142.
- Парра-Сааведра М., Гомес Л., Барреро А., Парра Дж., Вергара Ф., Наварро Э .: Прогнозирование преждевременных родов с использованием индекса консистенции шейки матки.Ультразвуковой акушерский гинеколь 2011; 38: 44-51.
- Бадир С., Мазза Э, Циммерманн Р., Байка М.: Смягчение шейки матки происходит на ранних сроках беременности: характеристика жесткости шейки матки у 100 здоровых женщин с использованием метода аспирации. Prenat Diagn 2013; 33: 737-741.
- Афзали Н., Мохаджери М., Малек А., Аламатин А. Область шейных желез: новый сонографический маркер в прогнозировании преждевременных родов.Arch Gynecol Obstet 2012; 285: 255-258.
- Kahyaoglu S, Kahyaoglu I, Kaymak O, Sagnic S, Mollamahmutoglu L, Danisman N: Может ли трансвагинальное ультразвуковое исследование эндоцервикальной железистой области предсказать преждевременные роды у пациентов, получавших токолитическую терапию по поводу угрожающих родов: кросс-секционное исследование.J Matern Fetal Neonatal Med 2013; 26: 920-925.
Автор Контакты
Нурия Баньос, доктор медицины, BCNatal — Барселонский центр материнско-фетальной медицины
и неонатальная медицина, Клиническая больница и больница Сан-Хуан-де-Деу
Fetal i + D Fetal Medicine Research Center, IDIBAPS
Барселонский университет, Сабино-де-Арана 1 , ES-08028 Барселона (Испания)
Эл. Почта NBANOS @ clinic.ub.es
Подробности статьи / публикации
Предварительный просмотр первой страницы
Получено: 15 января 2015 г.
Принято: 8 мая 2015 г.
Опубликовано в Интернете: 26 июня 2015 г.
Дата выпуска: ноябрь 2015 г.
Количество страниц для печати: 9
Количество рисунков: 1
Количество столов: 4
ISSN: 1015-3837 (печатный)
eISSN: 1421-9964 (онлайн)
Для дополнительной информации: https: // www.karger.com/FDT
Авторские права / Дозировка препарата / Заявление об ограничении ответственности
Авторские права: Все права защищены. Никакая часть данной публикации не может быть переведена на другие языки, воспроизведена или использована в любой форме и любыми средствами, электронными или механическими, включая фотокопирование, запись, микрокопирование, или какой-либо системой хранения и поиска информации, без письменного разрешения издателя. .
Дозировка лекарства: авторы и издатель приложили все усилия, чтобы гарантировать, что выбор и дозировка лекарства, указанные в этом тексте, соответствуют текущим рекомендациям и практике на момент публикации.Тем не менее, ввиду продолжающихся исследований, изменений в правительственных постановлениях и постоянного потока информации, касающейся лекарственной терапии и реакций на них, читателю рекомендуется проверять листок-вкладыш для каждого препарата на предмет любых изменений показаний и дозировки, а также дополнительных предупреждений. и меры предосторожности. Это особенно важно, когда рекомендованным агентом является новое и / или редко применяемое лекарство.
Отказ от ответственности: утверждения, мнения и данные, содержащиеся в этой публикации, принадлежат исключительно отдельным авторам и соавторам, а не издателям и редакторам.Появление в публикации рекламы и / или ссылок на продукты не является гарантией, одобрением или одобрением рекламируемых продуктов или услуг или их эффективности, качества или безопасности. Издатель и редактор (-ы) не несут ответственности за любой ущерб, причиненный людям или имуществу в результате любых идей, методов, инструкций или продуктов, упомянутых в контенте или рекламе.
Побуждающий труд
Что такое побуждающий труд?
Стимулирование родов (также называемое индукцией родов) — это когда ваш лечащий врач дает вам лекарство или использует другие методы, такие как взбивание воды (амниотический мешок), чтобы начать роды.Амниотический мешок (также называемый мешком с водой) — это мешок внутри матки (матки), в котором находится растущий ребенок. Мешочек наполнен околоплодными водами. Сокращения — это когда мышцы матки напрягаются, а затем расслабляются. Сокращения помогают вытолкнуть ребенка из матки.
Ваш поставщик медицинских услуг может порекомендовать стимулирование родов, если ваше здоровье или здоровье вашего ребенка находятся под угрозой, или если вы на 2 недели или более просрочены в срок. Для некоторых женщин стимулирование родов — лучший способ сохранить здоровье мамы и ребенка.
Если есть медицинские причины для того, чтобы назначить индукцию, поговорите со своим врачом о том, чтобы подождать, по крайней мере, до 39 недель беременности. Это дает вашему ребенку время, необходимое ему для роста и развития до рождения. Стимулирование родов должно производиться только по медицинским показаниям.
Как вызываются роды?
Ваш лечащий врач использует один или несколько из этих методов для стимулирования родов:
- Отделение амниотического мешка от стенки матки (также называется очисткой или очисткой плодных оболочек). Ваш врач осторожно проводит пальцем в перчатке через шейку матки и отделяет амниотический мешок от матки. Шейка матки — это вход в матку, который находится в верхней части влагалища. Вы можете сделать эту процедуру в офисе вашего провайдера. У вас могут быть спазмы или кровянистые выделения.
- Созревание шейки матки. Ваш врач дает вам лекарство под названием простагландины, которое помогает смягчить и истончить шейку матки, чтобы она открывалась во время родов. Вы можете принимать лекарство внутрь или вводить его во влагалище.Вы получаете лекарство в больнице. Ваш врач также может использовать во влагалище лекарство под названием ламинария. Он впитывает влагу и расширяется, открывая шейку матки. Или ваш провайдер может использовать инструмент, называемый лампой Фолея. Это тонкая трубочка с воздушным шариком на конце. Ваш врач вводит его во влагалище, чтобы расширить шейку матки.
- Давать лекарства от схваток. Медработники часто используют лекарство под названием окситоцин для стимуляции родов. Это лекарство представляет собой искусственную форму гормона, который помогает запускать схватки.В больнице ваш врач вводит окситоцин через капельницу (игла в вену). Это может вызвать сильные схватки. Спросите своего врача о обезболивающих, которые вам могут понадобиться во время родов.
- Разрыв воды (также называемый разрывом плодных оболочек или амниотомией). Ваш врач использует небольшой крючок, похожий на вязальную спицу, чтобы сломать амниотический мешок, на котором находится ваш ребенок. Это не должно быть болезненным, но вы можете почувствовать прилив теплой жидкости.
Направление родов может занять от нескольких часов до нескольких дней.Это зависит от того, как ваше тело реагирует на лечение.
Каковы риски принуждения к труду?
Риски включают:
- Ваш срок может быть не совсем правильным. Иногда бывает трудно точно определить, когда вы забеременели. Если вы назначили индукцию и у вас не установлен срок родов, ваш ребенок может родиться слишком рано. Если ваша беременность протекает нормально, подождите, пока роды начнутся сами по себе. Если вам необходимо назначить индукционный прием по медицинским показаниям, спросите своего поставщика, можете ли вы подождать не менее 39 недель.
- Окситоцин и лекарства, вызывающие созревание шейки матки, могут вызвать слишком тесные схватки. Это может снизить частоту сердечных сокращений вашего ребенка. Ваш врач внимательно следит за частотой сердечных сокращений вашего ребенка во время родов. Если частота пульса вашего ребенка изменится, ваш врач может прекратить или уменьшить количество принимаемых вами лекарств.
- Вы и ваш ребенок подвержены повышенному риску заражения. Амниотический мешок обычно защищает вашего ребенка и вашу матку от инфекции.Если после разрыва плодных оболочек роды начинаются через некоторое время, вероятность заражения возрастает.
- Возможно, проблема с пуповиной. Если амниотический мешок разорван, пуповина может проскользнуть во влагалище раньше, чем это сделает ваш ребенок. Это называется пролапсом пуповины. Это более вероятно, если у вашего ребенка тазовое предлежание. Это когда ягодица или ступни вашего ребенка перед рождением смотрят вниз, а не головой. Выпадение пуповины может вызвать сдавливание пуповины во время родов.В этом случае ваш ребенок не получает достаточно кислорода, что может быть опасно для жизни.
- Индукция может не работать, поэтому вам может потребоваться кесарево сечение (также называемое кесаревым сечением). Кесарево сечение — это операция, при которой ваш ребенок рождается через разрез, сделанный вашим врачом на животе и в матке.
- У вас может быть разрыв матки. Это когда матка рвется во время родов. Это случается редко, но может вызвать серьезное кровотечение. Если у вас было кесарево сечение во время предыдущей беременности, у вас повышенный риск разрыва матки, потому что после кесарева сечения в матке остается рубец.
- У вас может быть повышенный риск серьезного кровотечения после родов (так называемого послеродового кровотечения). Стимуляция родов увеличивает вероятность того, что мышцы матки не будут сокращаться должным образом после родов, что может привести к кровотечению.
Последнее обновление: сентябрь 2018 г.
См. Также: 39 недель инфографики
Frontiers | Индукция Agarwood: текущие разработки и перспективы на будущее
Введение
Agarwood (также известное как gaharu в Юго-Восточной Азии, oud на Ближнем Востоке, chen xiang в Китае, jinkoh в Японии и agar в Индии) представляет собой очень ценную ароматную темную смолистую сердцевину. из Aquilaria видов (Liu Y.Ю. и др., 2017). Образование агарового дерева обычно связано с ранением и грибковой инфекцией деревьев Aquilaria (Liu Y. et al., 2013; Mohamed et al., 2014). Смола выделяется деревьями в качестве защитной реакции и откладывается вокруг ран в течение нескольких лет после травмы, где накопление летучих соединений в конечном итоге приводит к образованию агарового дерева (Subasinghe and Hettiarachchi, 2013).
Древесина агара широко использовалась в качестве лечебных духов, народной медицины, в религиозных целях и в качестве ароматного пищевого ингредиента (Liu Y.и др., 2013). Некоторые из самых ранних известных применений агарового дерева были описаны в древней литературе, религиозных писаниях и медицинских текстах. Слово «алоэ», означающее «дерево агар», было обнаружено у санскритского поэта Калидаса, которое можно датировать ок. IV – V века н.э. (Ли и Мохамед, 2016). Между тем, использование агарового дерева в рецептах традиционной китайской медицины того же периода также было зарегистрировано. Китайская медицина использует его как натуральное успокаивающее, болеутоляющее, пищеварительное и ветрогонное средство (Ye et al., 2016; Лю Ю. Ю. и др., 2017).
Древесина агара пользуется большим спросом во всем мире как сырье для благовоний, парфюмерии и медицины, причем двумя основными регионами потребления являются Ближний Восток и Восточная Азия (Antonopoulou et al., 2010). По мере того как в последние десятилетия благосостояние стран-потребителей постепенно увеличивалось, рыночный спрос на агаровую древесину начал превышать ее предложение. Мировые цены на агаровую древесину могут варьироваться от 20 до 6000 долларов США за килограмм древесной щепы в зависимости от ее качества или 10 000 долларов США за килограмм самой древесины (Abdin, 2014).Кроме того, стоимость эфирного масла агарового дерева может достигать 30 000 долларов США за килограмм. Ежегодный мировой рынок агаровой древесины оценивается в диапазоне от 6 до 8 миллиардов долларов США (Akter et al., 2013), однако большое количество сделок не было зарегистрировано.
Aquilaria принадлежит к семейству покрытосеменных Thymelaeaceae, которое является эндемиком Индомалайского царства. На сегодняшний день зарегистрировано в общей сложности 21 видов Aquilaria, и 13 из них признаны производящими агаровую древесину (Lee and Mohamed, 2016).Однако разрушительная эксплуатация агарового дерева сильно повлияла на дикую популяцию всех видов Aquilaria . Как следствие, этот род в настоящее время внесен в список исчезающих видов и охраняется в соответствии с положениями Конвенции о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения (СИТЕС), из-за резкого сокращения численности видов в дикой природе (Конвенция о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения). [СИТЕС], 2004 г .; Ли и Мохамед, 2016 г.). Высокий спрос на качественную древесину агара в сочетании с истощением диких деревьев Aquilaria означал, что цена на древесину агара будет продолжать расти.В качестве альтернативы, массовое выращивание и большие плантации деревьев аквиларии , которые служат устойчивым источником для получения агаровой древесины, в значительной степени решили проблему нехватки агаровой древесины на мировом рынке.
Поскольку здоровое дерево Aquilaria не образует древесину агара, поэтому нехватка природного дерева агара побудила к разработке методов искусственного получения древесины агара. Попытки искусственно вызвать образование агарового дерева можно проследить еще до 300 C.E. в китайской истории, где было зарегистрировано, что отложение смолы, сопровождающееся изменением цвета внутренних тканей, может произойти в течение года в результате повреждения деревьев (López-Sampson and Page, 2018). Помимо механического ранения, в настоящее время в деревообрабатывающей промышленности все чаще используются химические методы, методы воздействия насекомых и патогенов (Liu Y. et al., 2013; Mohamed et al., 2014; Kalita, 2015). Все эти методы индукции в любом случае имитируют естественные процессы образования агарового дерева, которые имеют свои сильные и слабые стороны.В этой статье мы стремимся более полно охватить существующие методы индукции и перспективы их развития с использованием достижений биотехнологии. Чтобы лучше понять процесс образования агарового дерева, также будет разработан молекулярный механизм путей биосинтеза вторичных метаболитов, лежащих в основе производства смолы.
Подходы к индукции Agarwood
Беспорядочная заготовка агарового дерева в естественных средах обитания серьезно затруднила естественное возобновление деревьев аквиларии , тем самым поставив под угрозу выживание этого вида в дикой природе.Чтобы удовлетворить высокий рыночный спрос, но при этом защитить этот вид от исчезновения, в азиатских странах были созданы массовые плантации из деревьев Aquilaria , чтобы обеспечить устойчивое производство агаровой древесины (Azren et al., 2018). Поскольку формирование агаровой древесины в естественной среде — это очень длительный процесс, который может занять до 10 лет, разработке эффективной индукционной технологии было уделено большое внимание, поскольку чрезвычайно важно обеспечить стабильность урожайности агаровой древесины от одомашненных деревьев Aquilaria .
Естественно, образование агарового дерева часто связано с физическим ранением или повреждением деревьев Aquilaria в результате удара грома, выпаса животных, заражения вредителями и болезнями (Rasool and Mohamed, 2016; Wu et al., 2017). Эти события подвергают внутреннюю часть деревьев воздействию патогенных микробов, которые активируют защитный механизм Aquilaria , чтобы инициировать производство смолы. Этот естественный процесс формирования агарового дерева во многом вдохновил на разработку разнообразных методов искусственной индукции (Таблица 1).Например, многие традиционные методы индукции, такие как установка гвоздей, продырявление, сжигание, разрушение ствола и удаление коры, основаны на концепции физического ранения деревьев (Mohamed et al., 2010; Azren et al., 2018). Хотя это рентабельно и требует только персонала с небольшими научными знаниями или отсутствием научных знаний о древесине агара, эти методы индукции обычно приводят к низкому качеству и неопределенному выходу древесины агара.
Таблица 1. Сильные и слабые стороны различных методов индуцирования агарового дерева.
С более глубоким пониманием грибковых взаимодействий Aquilaria , способствующих формированию агарового дерева, методы индукции постепенно перешли от единственного механического ранения к преднамеренному нанесению биологического инокулята (Jong et al., 2014). Было обнаружено, что многие штаммы чистых культур грибов, выделенных из природного агарового дерева, являются эффективными биологическими агентами, вызывающими образование агарового дерева у здоровых деревьев Aquilaria (Cui et al., 2013; Siburian et al., 2015; Сангаресвари Нагаджоти и др., 2016). Сообщалось, что инфицированные грибком деревья Aquilaria откладывают смолу агарового дерева вокруг зараженных участков в качестве барьера для предотвращения дальнейшего проникновения грибов (Cui et al., 2013; Rasool and Mohamed, 2016). Одним из очевидных преимуществ использования грибкового инокулята является то, что он, как правило, считается безопасным в обращении и экологически чистым. Однако инокуляция грибов обычно приводит к локализованному и нестабильному качеству агарового дерева из-за различных используемых грибковых консорциумов.В качестве решения требуется трудоемкий процесс вырубки и длительное время инкубации, чтобы максимально увеличить площадь колонизированной поверхности на дереве для получения лучшего качества древесины агара (Mohamed et al., 2014).
Вместо того, чтобы полагаться на внешние стимулы для запуска реакции растений, будь то механическое ранение или биологический инокулят, были разработаны некоторые индукционные подходы для введения сигнальных молекул непосредственно и конкретно в деревья Aquilaria для инициирования путей биосинтеза смолы агарового дерева (Liu Y.et al., 2013; Wu et al., 2017). Химические индукторы обычно включают фитогормоны, соли, минералы и вещества биологического происхождения (Zhang et al., 2012; Liu Y. et al., 2013; Van Thanh et al., 2015). Кроме того, подходящий способ доставки часто разрабатывается вместе с химическими составами для облегчения крупномасштабного процесса индукции, например, сосуд, оборудованный иглой для переливания крови (Yang et al., 2014c). На сегодняшний день было разработано несколько подходов к индукции, основанных на концепции химической индукции, таких как набор для выращивания агаровой древесины (CA-kit), метод индукции агаровой древесины целого дерева (Agar-Wit) и метод биологической индукции агаровой древесины (Agar-bit).CA-kit — это комбинированный метод, основанный на физическом ранении и химической индукции, при котором индуцирующий агент вводится в дерево Aquilaria через устройство для аэрации, вставленное в рану (Blanchette and Heuveling, 2009). Этот метод дает удовлетворительный урожай и качество, но процедуры в некотором роде традиционны. С другой стороны, Agar-Wit — это химическая обработка с помощью транспирации для формирования общей раны на дереве, где предварительно загруженный индуктор в наборе для переливания распределяется через транспирацию растений (Liu Y.и др., 2013). С помощью этого метода может быть достигнута большая площадь покрытия агаровым деревом, но, к сожалению, образуются более разрушенные ткани. Точно так же метод Agar-bit использует идею распределения индуцирующего реагента путем транспирации растений, за исключением того, что реагенты вводятся непосредственно в стебли дерева (Wu et al., 2017).
Благодаря химической индукции можно свести к минимуму трудоемкий процесс высыхания, поскольку требуется меньше участков индукции для доставки индукторов по растениям посредством процесса транспирации.Было показано, что правильно подобранный индуктор может производить искусственную древесину агара с качеством, близким к качеству, полученному из природного источника (Liu Y. et al., 2013). Несмотря на быстрые результаты и высокие урожаи, применение химических индукторов по-прежнему вызывает скептицизм в отношении токсичности как для человека, так и для окружающей среды. Требуются дополнительные оценки химических индукторов, чтобы проверить их эффективность в полевых условиях, а также популяризировать их использование. Химические индукторы, несомненно, более подходят для массового производства с более легким контролем качества, чем биологический инокулят, который может заменить традиционные методы индукции и использование биологического инокулята в промышленности древесного агара.
Основные составляющие Agarwood
Главная привлекательность отрасли древесного агара — ее чрезвычайно высокая рыночная стоимость. Тем не менее, цена агарового дерева в значительной степени определяется его качеством, которое оценивается исключительно на основе человеческого опыта, основанного на многовековой практике каждой страны. Отсутствие стандартной системы оценки качества может быть связано с замысловатым внешним видом продаваемой древесины агара и личной заинтересованностью. Принятая в настоящее время оценка качества древесины агара на рынке была подробно рассмотрена Лю Ю.Y. et al. (2017). В последнее время все большее внимание уделяется анализу метаболитов агарового дерева, поскольку некоторые исследования показали, что существует корреляция качества агарового дерева с выходом его смолы и составляющими метаболитов (Pasaribu et al., 2015; Liu Y. Y. et al., 2017). Было проведено множество исследований для уточнения состава метаболитов агарового дерева, полученного либо дикими, либо искусственно индуцированными методами (Chen et al., 2012; Gao X. et al., 2014; Hashim et al., 2014). Был сделан вывод, что состав смолы агарового дерева в основном состоит из смесей сесквитерпенов и 2- (2-фенилэтил) хромонов (ПЭК) (Naef, 2011; Chen et al., 2012; Субасингхе и Хеттиараччи, 2015; Рисунок 1). Между тем было показано, что составляющими эфирного масла древесного агара являются в первую очередь сесквитерпеноиды (Fazila and Halim, 2012; Hashim et al., 2014; Jayachandran et al., 2014). Вместе все эти основные соединения и некоторые летучие ароматические метаболиты с низким содержанием летучих метаболитов образуют уникальное ароматное свойство древесины агара.
Рисунок 1. Основной молекулярный скелет сесквитерпенов (A) и 2- (2-фенилэтил) хромонов (B) .
Количество и типы составляющих метаболитов агарового дерева в каждом опубликованном исследовании варьируются в зависимости от источника агарового дерева, методов экстракции и используемых подходов к анализу (Fazila and Halim, 2012; Jong et al., 2014; Pasaribu et al., 2015). Тем не менее, согласно обзору Naef (2011), к настоящему времени в агаровой древесине из различных источников было идентифицировано более 150 соединений. Среди этих соединений 70 сесквитерпенов и около 40 типов РЕС, которые были распознаны в древесине агара, и их структура выяснена (Naef, 2011).Согласно данным различных исследований, несколько сесквитерпенов чаще присутствуют в древесине агара, включая ароматендрен, агароспирол, β-агарофуран, гуайол и (-) — аристолен (Fazila and Halim, 2012; Liu Y. et al., 2013; Jayachandran et al. ., 2014; Jong et al., 2014; рисунок 2). Сообщается, что некоторые сесквитерпены видоспецифичны, например, jinkoh-eremol и epi -γ-eudesmol, которые присутствуют только в A. malaccensis , тогда как баимуксинал существует только в A. crassna и A.sinensis (Naef, 2011; Liu Y. et al., 2013; Jong et al., 2014; Hashim et al., 2016). Стоит отметить, что в исследовании Pasaribu et al. (2015) было обнаружено, что содержание ароматендрена выше в агаровой древесине более высокого качества, и поэтому он был предложен в качестве эффективного химического маркера для сортировки агаровой древесины. Помимо аромадендрена, Jayachandran et al. (2014) позже предложили дополнительный маркерный валентен, который может иметь важное значение при классификации масла агарового дерева.
Рисунок 2. Химические структуры сесквитепеновых соединений, которые обычно присутствуют в смоле агарового дерева.
Производные PEC, как и другие основные ароматические компоненты агарового дерева, вносят важный вклад в сладкий, фруктовый и стойкий аромат агарового дерева при его сжигании. Эти соединения могут быть обнаружены только методами сверхкритической двуокиси углерода и экстракции растворителем, но никогда не присутствуют в экстракте гидродистилляции (Yoswathana, 2013; Jong et al., 2014). По сравнению с сесквитерпеновыми составляющими в древесине агара, типы РЕС, определяемые с помощью ГХ-МС, относительно ограничены.Структурные исследования показали, что все ранее описанные ПЭК в древесине агара имеют один и тот же основной скелет (молекулярная масса: 250) и аналогичные заместители, то есть либо гидрокси-, либо метоксигруппы (Mei et al., 2013). Процентное содержание 2 (2-фенилэтил) хромона и 2- (2-4-метоксифенилэтил) хромона в агаровой древесине высокого качества, такой как kanankoh , может достигать 66,47%, что в подавляющем большинстве случаев выше, чем в древесине более низкого качества. agarwood jinkoh , в котором всего 1,5% (Ishihara et al., 1993). Кроме того, наличие определенных производных РЕС в древесине агара было предложено как полезное при оценке классификации продуктов из древесины агара (Shimada et al., 1982). Существует 17 типов производных хромона, которые являются специфичными для агарового дерева и являются потенциальными маркерами для аутентификации (Naef, 2011). Было показано, что замещенные хромоны, такие как агаротетрол и изоагаротетрол (рис.3), имеют положительную корреляцию с качеством агаровой древесины, полученной на рынке, за некоторыми исключениями (Shimada et al., 1986).
Рис. 3. Химическая структура производных 2- (2-фенилэтил) хромона, обычно присутствующих в смоле агарового дерева.
Типы и производные основных соединений в древесине агара чрезвычайно широки и разнообразны, что указывает на различные ароматические свойства древесины агара из разных видов и региональных источников. Лучшее понимание метаболитов агарового дерева определенно облегчит идентификацию общепринятых биомаркеров для классификации агарового дерева.После публикации всеобъемлющего обзора Naef (2011), посвященного основным составляющим агарового дерева, новые соединения продолжают обнаруживаться в более поздних исследованиях (Wu et al., 2012a; Yang et al., 2014b; Wang et al., 2015 ). Количество обнаруженных соединений в древесине агара в будущем, безусловно, будет увеличиваться.
Пути биосинтеза составляющих древесины агара
ОбразованиеAgarwood может быть связано с механизмом самозащиты деревьев Aquilaria в ответ на биотические и абиотические стрессы (Gao et al., 2012b; Сингх и Шарма, 2015). Стресс вызывает защитные реакции видов Aquilaria , которые, в свою очередь, инициируют биосинтез вторичных метаболитов и накопление смолы агарового дерева. Ранее мы упоминали, что сесквитерпены и производные РЕС являются основными составляющими древесного агара. Следовательно, крайне важно понять метаболический путь регуляции и биосинтеза сесквитерпенов и производных хромона у видов Aquilaria , чтобы эффективно индуцировать образование агарового дерева.
Считается, что в растениях предшественники изопреноидов для биосинтеза сесквитерпенов, тритерпенов и стеринов образуются в цитозоле путем мевалоновой кислоты (MVA). В пластидах 1-дезокси-D-ксилулозо-5-фосфатный (DXP) или известный как метилэритритолфосфатный (MEP) путь обеспечивает прекурсоры для производства монотерпенов, дитерпенов и каротиноидов (Rohmer, 1999; Dong et al., 2015 ; Сингх и Шарма, 2015). Эти два пути биосинтезируют предшественник гомоаллильного изопреноида C5, то есть изопентенилпирофосфат (IPP) и его электрофильный аллильный изомер диметилаллилпирофосфат (DMAPP).Обмен IPP и DMAPP наблюдался между пластидами и цитозолем даже при пространственном разделении двух путей (Dong et al., 2015). Производство предшественников IPP и DMAPP из пирувата и ацетил-КоА включает ряд ферментов согласно соответствующему пути (рис. 4). Гены, кодирующие эти ферменты, были идентифицированы у видов Aquilaria с помощью анализа секвенирования транскриптома (Xu et al., 2013; Ye et al., 2016). Эти изопреновые звенья C5 позже будут направлены на образование фарнезилпирофосфата C15 (FPP) посредством последовательных реакций конденсации в присутствии FPP-синтазы (FPS) (Rohmer, 1999; Yang et al., 2013; Ye et al., 2016). FPS является одним из ключевых ограничивающих ферментов, ответственных за биосинтез сесквитерпена (Gaffe et al., 2000; Yang et al., 2013; Liu X. M. et al., 2017). Гены, кодирующие FPS, были клонированы из Aquilaria microcarpa ( Am-FaPS-1 ) (Kenmotsu et al., 2011) и Aquilaria sinensis ( AsFPS1 ) (Yang et al., 2013). Сообщалось, что уровень транскрипта AsFPS1 выше в стебле и корнях, чем в листьях, что позволяет предположить, что синтез сесквитерпена у видов Aquilaria имеет тенденцию к тканеспецифичности.Кроме того, было показано, что экспрессия Am-FaPS-1 повышается при воздействии метилжасмоната (MeJA), дрожжевого экстракта и Ca 2+ -ионофора A23187, что указывает на то, что два первых химических вещества эффективны для инициирования путь биосинтеза сесквитерпена, тогда как Ca 2+ может действовать как сигнальная молекула во время процесса активации (Kenmotsu et al., 2011). Это дает ключ к разгадке искусственной индукции образования агарового дерева с помощью экзогенных химически индуцированных подходов, запускающих путь биосинтеза сесквитерпена в деревьях Aquilaria .
Рисунок 4. Схематическая взаимосвязь между индуцированными раной механизмами передачи сигнала для биосинтеза сесквитерпена и регуляцией у видов Aquilaria для производства агарового дерева. Внешние стимулы запускают сигнальный путь Ca 2+ и вызывают защитные реакции видов Aquilaria через путь перекиси водорода (H 2 O 2 ), сигналы этилена (ET), сигналы жасмоновой кислоты (JA) и сигналы салициловой кислоты (SA).Лечение MeJA запускает продукцию H 2 O 2 , которая может вызывать запрограммированную гибель клеток (PCD) и увеличивать синтез сесквитерпена. Эти сигнальные молекулы активируют факторы транскрипции, такие как MYB, MYC и WRKY, которые будут связываться с цис--элементом на промоторе генов биосинтеза терпенов в путях мевалоновой кислоты (MVA) и метилэритритолфосфата (MEP), а также нижестоящие гены терпен-синтазы ( TP s). Прямые и косвенные взаимодействия показаны сплошными и пунктирными линиями соответственно. AACT , ацетил-CoA C-ацетилтрансфераза; HMGS , гидроксиметилглутарил (HMG) -CoA-синтаза; HMGR , HMG-CoA редуктаза; MK , мевалонаткиназа; MPK , фосфомевалонаткиназа; MDD , мевалонатдифосфатдекарбоксилаза; DXP , 1-дезокси-D-ксилулоза 5-фосфат; DXS , DXP-синтаза; DXR , редуктоизомераза DXP; CMK , 4- (цитидин 50-дифосфо) -2-C-метил-D-эритритолкиназа; MCS , 2-C-метил-D-D-эритритол-2,4-цикло-дифосфатсинтаза; HDS , (E) -4-гидрокси-3-метилбут-2-енилдифосфатсинтаза; IDS , изопентенилдифосфатсинтаза; GPS , геранилдифосфатсинтаза; GGPS , геранилгеранилдифосфатсинтаза.
На заключительной стадии производства сесквитерпенов ферменты, ответственные за диверсификацию сесквитерпена, в основном происходят из классов сесквитерпенсинтаз (SesTPs) и цитохром P450-зависимых монооксигеназ (P450s). Ферменты SesTP ответственны за катализирование образования полициклических каркасных комплексов из FPP с последующей окислительной функционализацией полученных каркасов ферментами цитохрома P450. Добавленные P450 гидроксильные группы могут служить в качестве молекулярных ручек для дальнейших модификаций, таких как алкилирование, этерификация и добавление сахарных остатков (Pateraki et al., 2015). Кроме того, ферменты P450, которые осуществляют стереоспецифическое гидроксилирование углеводородных основных цепей, что важно для новых хиральностей и дальнейших модификаций молекул сесквитерпена, до сих пор не были обнаружены у видов Aquilaria . Аналогичным образом, НАДФН-зависимые оксидоредуктазы цитохрома P450 (POR) в Aquilaria , которые действуют как окислительно-восстановительные партнеры активности катализа P450, в основном не исследованы. Сообщалось о нескольких исследованиях по выделению генов, кодирующих SesTP, из Aquilaria , что можно рассматривать как ранние попытки изучить SesTP, участвующие в формировании агарового дерева (Kumeta and Ito, 2010; Xu et al., 2013). В исследовании Kumeta and Ito (2010) пять генов, кодирующих сесквитерпенсинтазы, которые имеют очень похожие аминокислотные последовательности, были выделены из Aquilaria crassna . Три из этих генов были успешно экспрессированы в Escherichia coli и ферментативно преобразовали FPP в δ-гвайен в качестве своего основного продукта. Кроме того, три других гена сесквитерпенсинтазы ( ASS1 , ASS2 и ASS3 ), идентифицированные из A.sinensis посредством секвенирования транскриптомов, как было установлено, кодирует ферменты, которые также продуцируют δ-гвайен (Xu et al., 2013). Выделение генов SesTP было также описано в A. malaccensis , где временная и пространственная экспрессия двух SesTP s, о которых сообщалось в исследовании, то есть гуайена ( AmGuaiS1 ) и сесквитерпенсинтазы ( AmSesTPS1 ), было выяснено (Azzarina et al., 2016). Было обнаружено, что AmSesTPS1 высоко экспрессируется через 6 часов ранения, в то время как AmGuaiS1 индуцируется через 2 часа ранения с величиной 18 и 5.Соответственно в 5 раз выше, чем у неповрежденного контроля. Недавно новый ген сесквитерпенсинтазы ( As-sesTPS ) был выделен из A. sinensis , где рекомбинантный As-sesTPS катализировал FPP в неролидол (Ye et al., 2018). Анализ экспрессии показал, что уровень транскрипта As-sesTPS был намного выше в древесине агара, чем в здоровой древесине, подразумевая, что ген может участвовать в образовании древесины агара. Несмотря на то, что многие сесквитерпеновые соединения были обнаружены в древесине агара, соответствующие SesTP, ответственные за их производство, еще не сообщены из Aquilaria .По этой причине пути биосинтеза сесквитерпена участвуют в функционализации терпенов в . Aquilaria требует дальнейшего разъяснения.
С другой стороны, хромоны представляют собой большую группу вторичных метаболитов с широким спектром потенциальных терапевтических показаний в отношении иммуномодуляции, воспаления, рака, диабета, неврологических состояний, бактериальных и вирусных инфекций (Khadem and Marles, 2011; Yang et al., 2012 ; Tawfik et al., 2014). Хромон является производным полициклического органического соединения, а именно бензопиранового кольца, с замещением кетогруппой в его оксимном кольце.Обычно считается, что образование хромонов происходит как следствие конвергенции множественных путей биосинтеза вторичных метаболитов, включая путь пентакетида, путь шикимовой кислоты и добавление азотистого фрагмента из аминокислот или других источников (Khadem and Marles, 2011). Благодаря обширным фармакологическим свойствам, связанным с его бициклической кольцевой структурой, хромоны использовались в качестве привилегированной основы при разработке новых лекарств (Reis et al., 2017). PEC представляют собой небольшой класс хромонов, которые содержат фенилэтил-заместитель у C 2 бензопиранового кольца хромона, который оказался структурно уникальным в семействе (Ibrahim and Mohamed, 2015). До сих пор было обнаружено, что РЕС присутствуют только у нескольких видов растений, например Bothriochloa ischaemum (Wang et al., 2001), Imperata cylindrical (Liu X. et al., 2013), Cucumis melo L. (Ibrahim, 2014), Gyrinops salicifolia (Shao et al., 2016) и видов Aquilaria (Wu et al., 2012b; Yang et al., 2014a). Недавно Liao et al. Предложили гипотетическую схему пути биосинтеза ПЭК. (2018) на основе глубокого анализа химических компонентов древесины агара с использованием методов GC-EL-MS и UPLC-ESI-MS / MS. В их исследовании было обнаружено, что PEC являются основными компонентами смолы агарового дерева, которая состоит в основном из 2- (2-фенилэтил) хромонов типа флиндерсии (FTPEC). Далее выясняется, что образование FTPEC может катализироваться поликетидсинтазой (PK) типа III путем конденсации аналогов дигидро-циннамоил-КоА и малонил-КоА с 2-гидроксибензоил-КоА с образованием каркаса PEC, который впоследствии будет катализироваться гидроксилазы или O -метилтрансферазы (OMT) с образованием структурно различных FTPEC (Liao et al., 2018). Недавнее исследование показало, что солевой стресс может вызывать биосинтез ПЭК в каллусах A. sinensis (Wang et al., 2016). Транскриптомный анализ этих индуцированных солей каллусов A. sinensis выявил несколько активированных генов-кандидатов, потенциально участвующих в биосинтезе РЕС, включая три гена, кодирующих ОМТ (флавонол-ОМТ 1, флавонол-3-ОМТ и кофеил-КоА-ОМТ. ), а ген поликетидсинтазы III типа кодирует халконсинтазу 1 (AsCHS1).
Несмотря на недавний прогресс, достигнутый в понимании биосинтеза PEC, необходимы огромные усилия, чтобы экспериментально определить недостающие шаги в этом сложном пути биосинтеза PEC.Даже при недостаточных знаниях о детальном пути биосинтеза PEC искусственный синтез хромонов и некоторых их производных, тем не менее, возможен благодаря развитию химических процессов (Goel and Makrandi, 2006; Tawfik et al., 2014). Древесина агара — богатый источник производных ПЭК, который заслуживает дальнейшего изучения, чтобы раскрыть структуру новых хромоновых соединений и улучшить понимание механизма его биосинтеза на молекулярном уровне.
Сигнальный и регулирующий механизм образования Agarwood
В природе вероятность получения деревьев Aquilaria , содержащих агаровую древесину, чрезвычайно мала (1-2%), тогда как их можно найти только на патогенно инфицированных или раненых деревьях (Cui et al., 2013; Чипа, Кошик, 2017). Следовательно, разумно предположить, что существует индуцируемый раной процесс передачи сигнала, вызывающий экспрессию сесквитерпенсинтаз до образования агарового дерева. Чтобы прояснить взаимосвязь между передачей раневого сигнала и регуляцией образования агарового дерева, в последнее время внимание исследователей привлекли высокопроизводительные исследования образования агарового дерева (таблица 2). Схематическая диаграмма предлагаемого механизма передачи сигнала биосинтеза и регуляции сесквитерпена у видов Aquilaria представлена на рисунке 4.
Таблица 2. Избранные публикации по передаче сигналов и регуляции образования агарового дерева.
Сигнальный путь митоген-активируемой протеинкиназы (MAPK) был предложен в качестве индуцированного раной сигнального механизма для образования агарового дерева в A. sinensis , который фосфорилирует нижестоящие факторы транскрипции (TF), такие как MYB или WRKY, которые в конечном итоге приводят к экспрессии генов сесквитерпенсинтазы ( ASS s) (Xu et al., 2013). Сигнальный каскад MAPK состоит из трех последовательно активируемых компонентов [MAPK-киназные киназы (MAPKKKs), MAPK-киназы (MAPKKs) и MAPKs], который является высококонсервативным сигнальным механизмом у эукариот, опосредующим внеклеточные сигналы нижестоящим чувствительным генам (Sinha et al., 2011; Xu C. et al., 2017). Xu et al. (2013) сообщили, что всего 41 униген из анализа транскриптома раненых A. sinensis аннотированы как связанные с сигнальным путем MAPK, а 25 — с сигнальными путями кальция, которые могут играть роль в индуцированном раной образовании агарового дерева. В растениях ионы кальция (Ca 2+ ) являются важными внутриклеточными вторичными молекулами-посредниками для регулирования многих путей передачи сигналов, реагирующих на внешние раздражители (Tuteja and Mahajan, 2007).Более ранние исследования на других растениях показали, что ТФ являются ключевыми регуляторами в чувствительных к стрессу сигнальных путях для передачи сигналов в различные клеточные центры для активации механизмов адаптации / защиты растений от неблагоприятных условий окружающей среды, включая такие ТФ, как bZIP, ERF, EIN3, MYB, MYC и WRKY (Ambawat et al., 2013; Phukan et al., 2016; Schmiesing et al., 2016). Было обнаружено, что сверхэкспрессия AaWRKY1 в Artemisia annua положительно регулирует экспрессию гена аморфо-4,11-диен-синтазы ( ADS ) и значительно увеличивает продукцию артемизинина (Ma et al., 2009). Экспрессия TF GaWRKY хлопка также активирует гены сесквитерпен-синтазы для биосинтеза (+) — δ-кадинена и госсипол-сесквитерпена (Xu et al., 2004).
Помимо механического ранения, MeJA является эффективным элиситором для увеличения содержания сесквитерпенов в Aquilaria (Xu et al., 2013, 2016; Xu Y. H. et al., 2017). Предыдущие исследования показали, что тепловой шок может увеличивать экспрессию генов, участвующих в биосинтезе жасмоновой кислоты (JA), включая алленоксидциклазу ( AOC ), алленоксидсинтазу ( AOS ), липоксигеназу ( LOX ) и 12 -оксофитодиеноатредуктазы 3 ( OPR3 ), которые впоследствии приводят к продукции JA и накоплению сесквитерпена в A.sinensis суспензионной культурой клеток (Xu et al., 2016). Коронатин-нечувствительный белок 1 (AsCOI1) A. sinensis , действующий как рецептор в сигнальном пути MeJA, был клонирован и охарактеризован (Liao et al., 2015). Исследование экспрессии AsCOI1 продемонстрировало, что ген экспрессируется по тканеспецифическому паттерну, который наиболее высок в стебле, за которым следуют корень и листья. Полученные данные предполагают, что производство смолы из-за внешних раздражителей может быть более чувствительным в стебле Aquilaria .Ген AsCOI1 реагирует на раннее лечение MeJA, механическое ранение и тепловой стресс. Применение MeJA в A. sinensis увеличило уровень экспрессии 17 генов, связанных с передачей сигналов из ран, включая TF WRKY4 и MYB4 , протеинкиназы CAPK s, MAPK s и MAPKP , NADKP , оксидаза , noxB и некоторые регуляторы, относящиеся к сигнальным молекулам MeJA, этилена и перекиси водорода (Xu et al., 2013). Интересно, что пероксид водорода (H 2 O 2 ), продуцирующий НАДФН-оксидазу noxB, был значительно активирован обработкой MeJA (Xu et al., 2013; Gong et al., 2017), что согласуется с с открытиями, что MeJA запускает продукцию H 2 O 2 в растениях (Orozco-Cardenas et al., 2001; Hung et al., 2006). Исследование суспензионных культур A. sinensis показало, что H 2 O 2 может вызывать запрограммированную гибель клеток (PCD) и синтез сесквитерпена за счет повышенной экспрессии генов ASS из-за эндогенного накопления салициловой кислоты. (SA) (Liu et al., 2015). Между тем, экспрессия чувствительной к жасмонату ключевой сесквитерпенсинтазы ASS1 в A. sinensis , как было описано, регулируется TF AsMYC2 (Xu Y. H. et al., 2017). В качестве гена немедленного раннего ответа на лечение MeJA, AsMYC2 связывается с промотором ASS1 , содержащим мотив G-бокса, чтобы инициировать экспрессию ASS1 . Аналогичным образом было показано, что гомолог AsMYC2 в Arabidopsis (MYC2) реагирует на MeJA и повышает экспрессию двух сесквитерпен-синтаз ( TPS11 и TPS21 ) при лечении (Hong et al., 2012). Кроме того, обработка MeJA успешно индуцировала синтез 3 сесквитерпенов, которые представляют собой α-гуайен, α-гумулен и δ-гвайен, в культуре клеток A. crassna (Ito et al., 2005; Kumeta and Ito, 2010).
В отличие от пути биосинтеза сесквитерпена, биосинтез и регуляция PECs остаются почти неизвестными. В исследовании Wang et al. (2016) сорок один РЕС был получен в каллусах A. sinensi обработкой солью. С помощью высокопроизводительного анализа транскриптомов в общей сложности 18 069 дифференциально экспрессируемых транскриптов между контролем и обработанными NaCl A.sinensis , индуцированные 24- или 120-часовым солевым стрессом. Сообщается, что многие дифференциально экспрессируемые гены участвуют в передаче гормонального сигнала; в том числе гены, кодирующие каскады MAPK, рецептор-подобные киназы, факторы трансдукции сигнала Ca 2+ и транскрипционные факторы (Wang et al., 2016).
Тем не менее, темпы исследований не только остановились на поиске эффективных индукторов и лежащего в их основе процесса передачи сигнала, но также распространились на их механизм посттранскрипционной регуляции.Путем глубокого секвенирования мРНК из здоровых контрольных и раненых образцов A. sinensis , Gao et al. (2012a) идентифицировали десять стресс-чувствительных miRNAs из 74 предполагаемых консервативных miRNAs, и их предшественники, образующие шпильки, также были подтверждены. Паттерн экспрессии показал, что шесть из этих стресс-зависимых miRNA были активированы, в том числе miR159, miR168, miR171, miR396, miR397 и miR408, тогда как miR160 и miR398 были подавлены и продолжали снижаться на 2-дневной точке (Gao et al. al., 2012а). Различные ответы miRNAs реагировали на лечение, и эффект длился в течение разного времени, отражая разнообразие их положений в посттранскрипционной регуляции раневого ответа у A. sinensis . Среди идентифицированных miRNAs интерес представляет подавляемая miR398, где было продемонстрировано, что она негативно регулирует вызываемое патогенами молекулярные паттерны (PAMP) отложения каллозы и врожденный иммунитет растений против бактерий (Li et al., 2010).Противоположно прореагировавшие miR160 и miR398 в A. sinensis предполагают, что они могут быть важными регуляторами и играть более определенную роль в формировании агарового дерева.
Дальнейшее исследование профилей miRNAs раненых A. sinensis показало, что некоторые из наиболее консервативных miRNA, такие как семейства miR159 и miR396, быстро увеличиваются и исчезают в раннем периоде лечения, что подразумевает их функцию в начале реакции на рану (Gao ZH et al. ., 2014). Подавление miR396b2 в поврежденных тканях A.sinensis , как предполагалось, участвует в биосинтезе и накоплении компонентов древесины агара. Считалось, что мишень miR396b2, которая обладает активностью глутамил-тРНК редуктазы, связывает НАДФ и продуцирует НАДФН. НАДФН является кофактором двух ключевых ферментов биосинтеза терпенов, а именно 1-дезокси-D-ксилулозо-5-фосфата (DXR) и 3-гидрокси-3-метилглутарил-КоА (HMGR) (Nagegowda, 2010). Кроме того, растительные P450, которые окислительно функционализируют терпеновые каркасы, также требуют восстановителей для своей каталитической активности, которая обычно обеспечивается NADH или NADPH.Несмотря на отсутствие секвенированного генома, высокопроизводительный анализ транскриптома обеспечивает реальный подход для изучения общих изменений экспрессии генов у видов Aquilaria , реагирующих на различные стрессы. Дальнейшее изучение функций идентифицированных регуляторных последовательностей поможет выявить механизм регуляции образования агарового дерева.
Будущие перспективы индукционной технологии Agarwood
Предыдущие исследования показали, что на формирование древесины агара могут влиять многие факторы.Считается, что вместе с чрезвычайно сложной композицией смолы агарового дерева образование агарового дерева представляет собой сложный процесс, который включает в себя множество физиологических изменений, происходящих на деревьях Aquilaria , чтобы справиться с внешними стимулами в форме биотических или абиотических. Весь этот процесс образования агарового дерева в любом случае неотделим от реакции деревьев на экспрессию генов на триггерные факторы. Следовательно, при дальнейшем усовершенствовании технологии индукции древесного агара следует акцентировать внимание на двух аспектах, направленных на дальнейшее повышение эффективности индукции и скрининг более чувствительных линий Aquilaria для производства смолы в рамках программы селекции.
Для повышения эффективности индукции решающую роль играет индукционный метод. Концепцию доступных в настоящее время индукционных подходов можно резюмировать как предоставление внешних стимулов для активации продукции сигнальных молекул растений, которые в конечном итоге приводят к биосинтезу смол, или обход внешних стимулов путем прямого введения сигнальных молекул растениям. В любом случае, общая проблема заключается в увеличении урожайности и качества древесины агара, а также в сокращении вмешательства человека (например,g., высверливание) во время индукционного процесса. Поскольку вышеупомянутый метод физического ранения и биологической индукции имеет свои неизбежные недостатки в виде нестабильного качества древесины агара и необходимости интенсивной рабочей силы, метод химической индукции можно рассматривать как многообещающий подход для дальнейшей оптимизации (Таблица 1). Всестороннее понимание образования агарового дерева на молекулярном уровне за счет высокой пропускной способности с использованием омиксного подхода, такого как транскриптомный и метаболомный, оказывается полезным для более целенаправленного и направленного улучшения индукционной рецептуры, а не экспериментов, основанных на пробах и ошибках.
Благодаря сочетанию с подходами omics, такими как технология секвенирования одной молекулы в реальном времени (SMRT), которая предлагает более длинные длины чтения и очень смежные сборки de novo (Rhoads and Au, 2015), она имеет тенденцию быть особенно полезной для нерешенных проблем в геноме. и транскриптом немодельных видов Aquilaria , геномы которых недоступны. При более длинных считываниях можно легко получить высокоповторяющуюся некодирующую регуляторную последовательность генов, контролирующих продукцию смолы агарового дерева.Интеграция на основе сходства / идентичности последовательностей может использоваться для установления наборов данных перекрестных ссылок между запрашиваемыми последовательностями и их гомологичными ссылками из различных общедоступных баз данных и генетических ресурсов, где могут быть использованы структуры геномных последовательностей, особенности доменов, промоторные области и генная онтология для мотивов. быть назначенным (Mochida and Shinozaki, 2011). Развитие технологии секвенирования сделало будущие исследования полногеномного секвенирования видов Aquilaria легче восполнить недостаток полногеномной информации в текущей ситуации.Интеграция данных на основе последовательности генома важна для анализа глобальных изменений транскриптома с помощью микрочипов всего генома. Анализ экспрессии генов индуцированной Aquilaria в целом может быть исследован в общих скоординированных тенденциях с помощью этого подхода, который невозможно различить в отдельных анализах. Профили экспрессии, полученные таким образом, помогут идентифицировать гены потенциальных биомаркеров, продуцирующих древесину агара, которые являются важными индикаторами для последующих применений индукции агарового дерева.
Интеграция подходов системной биологии и омики, охватывающих геномику, транскриптомику, протеомику, метаболомику и функциональный анализ; представляют собой потенциальное решение для понимания мультигенной природы биосинтеза смол в Aquilaria . На основе большого количества ранее проведенных экспериментов по индукции агарового дерева, транскриптомных исследований и исследований метаболитов (Naef, 2011; Gao X. et al., 2014; Ye et al., 2016; Wu et al., 2017), интегрированный омикс-анализ может действительно служат платформой для построения более полной картины механизма биосинтеза смолы агарового дерева, который включает различные слои омика.Развитие высокопроизводительной масс-спектрометрии (МС), микрочипов и технологий секвенирования (ДНК и РНК) сделало возможным интегрировать такие данные в системную биологическую структуру с помощью интегрированных омиков, которые помогли бы предсказать взаимодействия ген-ген, идентифицировать драйвер гены и молекулярные признаки образования агарового дерева (рис. 5). Аналогичным образом, потенциальные сигнальные молекулы для образования агарового дерева могут быть предсказаны после резкого увеличения доступности данных. Экспериментально подтвержденные эффективные сигнальные молекулы могут быть добавлены в существующий рецепт химического индуктора для дальнейшего повышения эффективности индукции.Более того, предварительное определение состава смолы или качества древесины агара возможно при глубоком понимании ключевых показателей качества и конкретных путей, задействованных в производстве древесины агара, путем интегрированного выяснения различных профилей метаболитов и транскриптов при различных методах индукции. Дальнейшее повышение эффективности индукции древесного агара должно сопровождаться разработкой системы мониторинга для раннего обнаружения нереагирующих деревьев, которая могла бы избежать вырубки деревьев Aquilaria , которые были безуспешно индуцированы.Это может быть достигнуто путем мониторинга экспрессии набора генов, участвующих в биосинтезе смолы агарового дерева.
Рисунок 5. Схематическое изображение применения омических подходов в разработке индукторов агарового дерева. PacBio, Pacific Biosciences; Illumina, секвенирование Illumina; Пиросек, пиросеквенирование; RNAseq — секвенирование РНК; 2D-PAGE, 2-мерный электрофорез в полиакриламидном геле; DIGE, дифференциальный гель-электрофорез; ChIP seq, секвенирование иммунопреципитации хроматина; ГХ-МС, газовая хроматография-масс-спектрометрия; ЖХ-МС, жидкостная хроматография-масс-спектрометрия; ЯМР, ядерный магнитный резонанс.
Помимо эффективного индуктора, реакция деревьев Aquilaria на стимуляцию является еще одним определяющим фактором для производства древесины агара. Зная, что степень реакции растений на стимулы в значительной степени зависит от их генетического состава, ожидается, что использование высокочувствительной линии Aquilaria в качестве мишени для индукции приведет к дальнейшему увеличению урожайности агарового дерева, а не к оптимизации рецепта одного индуктора. Традиционно селективная селекция, основанная на фенотипической селекции, была принята для создания новых линий растений с желательными признаками.Современные технологии связывают эти полезные черты растений с генетическими (вариации ДНК / РНК) или биохимическими (сигнатуры метаболитов) маркерами, чтобы обеспечить селекцию с помощью маркеров (MAS). Подход MAS открывает большие перспективы для отбора элитных линий Aquilaria , поскольку эти биомаркеры можно применять для прогнозирования фенотипических характеристик до того, как эти особенности станут более заметными. Такие биомаркеры также можно использовать для разработки быстрых и целенаправленных диагностических тестов, которые помогут программе отбора.В качестве альтернативы для получения высокопродуктивной линии комбинированный подход генной инженерии (например, технология редактирования генома CRISPR-Cas9) с культурой ткани может предоставить возможность манипулировать ключевыми генами-регуляторами Aquilaria , участвующими в производстве агарового дерева, которые поможет точно настроить или перенаправить метаболический поток на желаемые метаболические пути. В целом, интегрированная и высокопроизводительная стратегия предоставит достаточно информации для постоянного улучшения методов индукции агарового дерева, что превосходит традиционный способ создания метода индукции, основанный на визуальном наблюдении и личном опыте.Более глубокое понимание основных соединений и механизма биосинтеза смолы агарового дерева значительно упростило бы контроль стабильности урожайности, качества и цены агаровой древесины в будущем.
Взносы авторов
CT, NI, II и ZZ внесли свой вклад в концепцию обзорной статьи. CT написал первый черновик и разделы рукописи. Все авторы внесли свой вклад в доработку рукописи, прочитали и одобрили представленную версию.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Авторы выражают благодарность Universiti Kebangsaan Malaysia за финансирование в рамках гранта MI-2018-009, предоставленного ZZ.
Сокращения
DMAPP, диметилаллилпирофосфат; DXP, 1-дезокси-D-ксилулозо-5-фосфат; FPP, фарнезилпирофосфат; FPS, FPP-синтаза; FTPEC, 2- (2-фенилэтил) хромон типа флиндерсии; HMGR, 3-гидрокси-3-метилглутарил-КоА; IPP, изопентенилпирофосфат; JA, жасмоновая кислота; MAPK, митоген-активированная протеинкиназа; MAS, селекция с помощью маркеров; MeJA, метилжасмонат; MEP, метилэритритолфосфат; MVA, мевалоновая кислота; НАДФН, дигидроникотинамид-адениндинуклеотидфосфат; OMT, O -метилтрансфераза; P450, цитохром P450-зависимая монооксигеназа; PAMP, молекулярный паттерн, связанный с патогенами; PCD, запрограммированная гибель клеток; ПЭК, 2- (2-фенилэтил) хромон; ПК, поликетидсинтазы; ПОР, НАДФН-зависимая оксидоредуктаза цитохрома Р450; SA, салициловая кислота; SesTP, сесквитерпен-синтаза; SMRT, технология секвенирования одиночных молекул в реальном времени; ТФ, фактор транскрипции.
Список литературы
Абдин, М. Дж. (2014). Древесная промышленность агара: в Бангладеш еще не используется. Внутр. J. Econ. Manag. Sci. 3, 163–166. DOI: 10.2139 / ssrn.2430055
CrossRef Полный текст | Google Scholar
Актер С., Ислам М. Т., Зулькефели М. и Хан С. И. (2013). Производство Agarwood — многопрофильная область, которую предстоит изучить в Бангладеш. Внутр. J. Pharm. Life Sci. 2, 22–32. DOI: 10.3329 / ijpls.v2i1.15132
CrossRef Полный текст | Google Scholar
Ambawat, S., Шарма П., Ядав Н. Р. и Ядав Р. К. (2013). Гены факторов транскрипции MYB как регуляторы ответов растений: обзор. Physiol. Мол. Биол. Растения. 19, 307–321. DOI: 10.1007 / s12298-013-0179-1.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Антонопулу, М., Комптон, Дж., Перри, Л. С., и Аль-Мубарак, Р. (2010). «Торговля и использование агарового дерева (уд) в Объединенных Арабских Эмиратах», в: TRAFFIC Юго-Восточная Азия. (Женева: секретариат СИТЕС).
Google Scholar
Азрен П. Д., Ли С. Ю., Эманг Д. и Мохамед Р. (2018). История и перспективы индукционной технологии производства агарового дерева из культивируемой аквиларии в Азии: обзор. J. For. Res. 30, 1–11. DOI: 10.1007 / s11676-018-0627-4.
CrossRef Полный текст | Google Scholar
Аззарина, А. Б., Мохамед, Р., Ли, С. Ю., и Назре, М. (2016). Временная и пространственная экспрессия генов терпенсинтазы, ассоциированная с образованием агарового дерева у Aquilaria malaccensis Lam. N. Z. J. For. Sci. 46:12. DOI: 10.1186 / s40490-016-0068-9.
CrossRef Полный текст | Google Scholar
Бланшетт Р. и Хёвелинг В. Б. Х. (2009). Культивируемое дерево агара. Патент США № 7638145. Миннесота: Университет Миннесоты.
Google Scholar
Chen, H.Q., Wei, J. H., Yang, J. S., Zhang, Z., Yang, Y., Gao, Z. H., et al. (2012). Химические компоненты древесины агара, происходящие из растений эндемичного рода Aquilaria. Chem.Биодайверы. 9, 236–250. DOI: 10.1002 / cbdv.201100077.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чипа, Х., Каушик, Н. (2017). Разнообразие грибов и бактерий, выделенных из дерева и почвы Aquilaria malaccensis , вызывает образование агароспирола в течение 3 месяцев после искусственного заражения. Фронт. Microbiol. 8: 1286. DOI: 10.3389 / fmicb.2017.01286.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Конвенция о международной торговле видами, находящимися под угрозой исчезновения [СИТЕС] (2004 г.).»Конвенция о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения. Рассмотрение предложений по внесению изменений в приложения-I и -II Aquilaria spp. и Gyrinops spp. »в Протоколах тринадцатого совещания Конференции Сторон. Бангкок.
Google Scholar
Цуй, Дж., Го, С., Фу, С., Сяо, П., и Ван, М. (2013). Влияние инокуляции грибов на образование коровьей древесины у Aquilaria sinensis . Подбородок. Sci.Бык. 58, 3280–3287. DOI: 10.1007 / s11434-013-5856-5.
CrossRef Полный текст | Google Scholar
Донг Л., Йонгедейк Э., Бауместер Х. и Ван Дер Крол А. (2015). Потенциал биосинтеза монотерпенов субклеточных компартментов растений. New Phytol. 209, 679–690. DOI: 10.1111 / Nph.13629.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фазила, К. Н., Халим, К. Х. К. (2012). Влияние замачивания на урожайность и качество масла агарового дерева. J. Trop. Для. Sci. 24, 557–564.
Google Scholar
Gaffe, J., Bru, J. P., Causse, M., Vidal, A., Stamitti-Bert, L., Carde, J. P., et al. (2000). LEFPS1, ген фарнезилпирофосфата томатов, высоко экспрессируемый во время раннего развития плодов. Plant Physiol. 123, 1351–1362. DOI: 10.1104 / стр.123.4.1351
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гао, X., Xie, M., Liu, S., Guo, X., Chen, X., Zhong, Z., et al. (2014).Хроматографический фингерпринт-анализ метаболитов в естественной и искусственной древесине агара с использованием газовой хроматографии-масс-спектрометрии в сочетании с хемометрическими методами. J. Chromatogr. Б Аналит. Technol. Биомед. Life Sci. 967, 264–273. DOI: 10.1016 / j.jchromb.2014.07.039.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гао, З. Х., Янг, Ю., Чжан, З., Чжао, В. Т., Мэн, Х., Цзинь, Ю. и др. (2014). Профилирование микроРНК при обработке ран в Aquilaria sinensis для выявления возможных микроРНК, участвующих в образовании агарового дерева. Внутр. J. Biol. Sci. 10, 500–510. DOI: 10.7150 / ijbs.8065.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гао, З. Х., Вэй, Дж. Х., Янг, Ю., Чжан, З., Сюн, Х. Я. и Чжао, В. Т. (2012a). Идентификация консервативных и новых микроРНК в Aquilaria sinensis на основе данных секвенирования малых РНК и последовательностей транскриптомов. Ген 505, 167–175. DOI: 10.1016 / j.gene.2012.03.072.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гао, З.Х., Вэй, Дж. Х., Ян, Ю., Чжан, З., и Чжао, В. Т. (2012b). Выбор и проверка эталонных генов для изучения связанного со стрессом образования агарового дерева Aquilaria sinensis . Plant Cell Rep. 31, 1759–1768. DOI: 10.1007 / s00299-012-1289-х.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гоэль, С.С., и Макранди, Дж. К. (2006). Синтез 2- (2-фенилэтил) хромонов. Indian J. Chem. 45Б, 535–536. DOI: 10.1002 / chin.200623140
CrossRef Полный текст | Google Scholar
Гонг, Б., Ян, Ю., Вэнь, Д., и Ши, К. (2017). Перекись водорода, продуцируемая НАДФН-оксидазой: новый нижестоящий сигнальный путь при толерантности к стрессу, вызванной мелатонином, у Solanum lycopersicum. Physiol. Растение. 160, 396–409. DOI: 10.1111 / ppl.12581.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хашим, Ю., Исмаил, Н., Аббас, П. (2014). Анализ химических соединений масла агарового дерева разных видов методом газовой хроматографии и масс-спектрометрии (ГХМС). IIUM Eng.J. 15, 55–60. DOI: 10.31436 / iiumej.v15i1.469.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хашим, Ю. З., Керр, П. Г., Аббас, П., и Мохд Салле, Х. (2016). Aquilaria spp. (агар) как источник полезных для здоровья соединений: обзор традиционного использования, фитохимии и фармакологии. J. Ethnopharmacol. 189, 331–360. DOI: 10.1016 / j.jep.2016.06.055.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хонг, Г.Дж., Сюэ, X. Y., Мао, Ю. Б., Ван, Л. Дж., И Чен, X. Y. (2012). Arabidopsis MYC2 взаимодействует с белками DELLA, регулируя экспрессию гена сесквитерпенсинтазы. Растительная клетка 24, 2635–2648. DOI: 10.1105 / tpc.112.098749.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хунг, К. Т., Сюй, Ю. Т., и Као, К. Х. (2006). Перекись водорода участвует в старении листьев риса, вызванном метилжасмонатом. Physiol. Растение. 127, 293–303. DOI: 10.1111 / j.1399-3054.2006.00662.x.
CrossRef Полный текст | Google Scholar
Ибрагим, С. Р., Мохамед, Г. А. (2015). Встречающиеся в природе 2- (2-фенилэтил) хромоны, выяснение структуры и биологическая активность. Нац. Prod. Res. 29, 1489–1520. DOI: 10.1080 / 14786419.2014.9
.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Исихара, М., Цунея, Т., и Унеяма, К. (1993). Компоненты летучего концентрата агарового дерева. J. Essent. Oil Res. 5: 3. DOI: 10.1080 / 10412905.1993.9698221.
CrossRef Полный текст | Google Scholar
Ито М., Окимото К. И., Ягура Т., Хонда Г., Киучи Ф. и Шимада Ю. (2005). Индукция продукции сесквитерпеноидов метилжасмонатом в суспензионной культуре клеток Aquilaria sinensis . J. Essent. Oil Res. 17, 175–180. DOI: 10.1080 / 10412905.2005.9698867
CrossRef Полный текст | Google Scholar
Джаячандран, К., Секар И., Партибан К. Т., Амиртам Д. и Суреш К. К. (2014). Анализ различных сортов масла агарового дерева ( Aquilaria malaccensis Lamk.) С помощью ГХ-МС. Indian J. Nat. Prod. Ресурс. 5, 44–47.
Google Scholar
Джонг П. Л., Паскаль Т. и Рози М. (2014). Газовая хроматография-масс-спектрометрический анализ экстрактов древесины агара из зрелых и молодых особей Aquilaria malaccensis . Внутр. J. Agric. Биол. 16, 644–648.
Google Scholar
Калита, Дж.(2015). Ассоциация Zeuzera conferta Walker по формированию агарового дерева у Aquilaria malaccensis Lamk. Asian J. Plant Sci. Res. 5, 4–9.
Google Scholar
Кенмоцу Ю., Огита С., Катох Ю., Ямамура Ю., Такао Ю., Тацуо Ю. и др. (2011). Метилжасмонат-индуцированное усиление активности экспрессии Am-FaPS-1, предполагаемого гена фарнезилдифосфатсинтазы из Aquilaria microcarpa . J. Nat. Med. 65, 194–197.DOI: 10.1007 / s11418-010-0451-4.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кумета Ю. и Ито М. (2010). Характеристика δ-гвайен-синтаз из культивируемых клеток Aquilaria, ответственных за образование сесквитерпенов в древесине агара. Plant Physiol. 154, 1998–2007. DOI: 10.1104 / стр.110.161828.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ли, С.Ю., и Мохамед, Р. (2016). «Происхождение и одомашнивание Aquilaria, важного рода, производящего древесину агара», в Agarwood: Science Behind the Fragrance , ed.Р. Мохамед. (Берлин: Springer Singapore), 1–20. DOI: 10.1007 / 978-981-10-0833-7_1
CrossRef Полный текст | Google Scholar
Ли Ю., Чжан К., Чжан Дж., Ву Л., Ци Ю. и Чжоу Дж. М. (2010). Идентификация микроРНК, участвующих в врожденном иммунитете растений, запускаемом молекулярными патогенами. Plant Physiol. 152, 2222–2231. DOI: 10.1104 / стр.109.151803.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ляо, Г., Дун, В.-H., Yang, J. -L., Li, W., Wang, J., Mei, W. -L., Et al. (2018). Мониторинг химического профиля образования агарового дерева в течение одного года и размышления о биосинтезе 2- (2-фенилэтил) хромонов. Молекулы 23: 1261. DOI: 10,3390 / молекулы23061261.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ляо, Ю., Вэй, Дж., Сюй, Ю., и Чжан, З. (2015). Клонирование, экспрессия и характеристика гена COI1 (AsCOI1) из Aquilaria sinensis (Lour.) Гилг. Acta. Pharm. Грех. Б. 5, 473–481. DOI: 10.1016 / j.apsb.2015.05.009.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лю Дж., Сюй Ю., Чжан З. и Вэй Дж. (2015). Перекись водорода способствует запрограммированной гибели клеток и накоплению салициловой кислоты во время индуцированной продукции сесквитерпенов в культивируемых суспензиях клеток Aquilaria sinensis . Функц. Plant Biol. 42, 337–346. DOI: 10.1071 / FP14189.
CrossRef Полный текст | Google Scholar
Лю, X.М., Тао, Т. Т., Мэн, X. X., Чжан, В. В., Чанг, Дж., И Сюй, Ф. (2017). Клонирование и анализ экспрессии гена фарнезилдифосфатсинтазы (FPPS) из Chamaemelum nobile. Нет. Бот. Хорти Агробо. Клуж-На. 45: 2. DOI: 10,15835 / NBHA45210858.
CrossRef Полный текст | Google Scholar
Лю X., Чжан Б.-Ф., Ян Л., Чжоу Г.-Х. и Ван З.-Т. (2013). Два новых хромона и новый флавоновый гликозид от imperata cylindrica. Подбородок. J. Nat. Med. 11, 77–80.DOI: 10.1016 / S1875-5364 (13) 60012-6.
CrossRef Полный текст | Google Scholar
Лю Ю., Чен Х., Ян Ю., Чжан З., Вэй Дж., Мэн Х. и др. (2013). Метод выращивания агаровой древесины из цельного дерева: новый эффективный метод получения высококачественной агаровой древесины на культивируемых деревьях Aquilaria sinensis . Молекулы 18, 3086–3106. DOI: 10,3390 / молекулы18033086.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лю, Ю.Я., Вэй, Дж. Х., Гао, З. Х., Чжан, З., и Лю, Дж. К. (2017). Обзор оценки качества и классификации древесины агара. Подбородок. Herb. Med. 9, 22–30. DOI: 10.1016 / S1674-6384 (17) 60072-8.
CrossRef Полный текст | Google Scholar
Лопес-Сэмпсон, А., Пейдж, Т. (2018). История использования и торговли агаровой древесиной. Экон. Бот. 72, 107–129. DOI: 10.1007 / s12231-018-9408-4.
CrossRef Полный текст | Google Scholar
Ма, Д., Пу, Г., Лей, К., Ма, Л., Wang, H., Guo, Y., et al. (2009). Выделение и характеристика AaWRKY1, фактора транскрипции Artemisia annua, который регулирует ген аморфо-4,11-диен-синтазы, ключевой ген биосинтеза артемизинина. Physiol растительных клеток. 50, 2146–2161. DOI: 10.1093 / pcp / pcp149.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Mei, W. -L., Yang, D. -L., Wang, H., Yang, J. -L., Zeng, Y. -B., Guo, Z. -K., Et al. (2013). Характеристика и определение 2- (2-фенилэтил) хромонов в древесине агара с помощью ГХ-МС. Молекулы 18, 12324–12345. DOI: 10,3390 / молекулы181012324.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мочида К. и Шинозаки К. (2011). Достижения в области омики и биоинформатики для системного анализа функций растений. Physiol растительных клеток. 52, 2017–2038. DOI: 10.1093 / pcp / pcr153.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мохамед Р., Джонг П. Л. и Камзиа А. К. (2014). Инокуляция грибами индуцирует агаровую древесину у молодых деревьев Aquilaria malaccensis в питомнике. J. For. Res. 25, 201–204. DOI: 10.1007 / s11676-013-0395-0.
CrossRef Полный текст | Google Scholar
Мохамед Р., Джонг П. Л. и Зали М. С. (2010). Разнообразие грибов в раненых стеблях Aquilaria malaccensis . Fungal Divers. 43, 67–74. DOI: 10.1007 / s13225-010-0039-z.
CrossRef Полный текст | Google Scholar
Наеф, Р. (2011). Летучие и полулетучие компоненты агарового дерева, зараженной сердцевины видов Aquilaria: обзор. Ароматизатор. J. 26, 73–87. DOI: 10.1002 / ffj.2034.
CrossRef Полный текст | Google Scholar
Nagegowda, D. A. (2010). Метаболизм летучих терпеноидов растений: гены биосинтеза, регуляция транскрипции и субклеточная компартментация. FEBS Lett. 584, 2965–2973. DOI: 10.1016 / j.febslet.2010.05.045.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ороско-Карденас, М. Л., Нарваес-Васкес, Дж., И Райан, К. А. (2001).Перекись водорода действует как вторичный посредник для индукции защитных генов у растений томата в ответ на ранение, системин и метилжасмонат. Растительная клетка 13, 179–191. DOI: 10.1105 / tpc.13.1.179
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пасарибу, Г. Т., Валуйо, Т. К., и Пари, Г. (2015). Анализ химических соединений, определяющих качество древесины агара. Индонезийский J. For. Res. 2: 7. DOI: 10.20886 / ijfr.2015.2.1.1-7
CrossRef Полный текст | Google Scholar
Патераки, И., Хескес, А., Хамбергер, Б. (2015). Цитохромы P450 для функционализации терпенов и метаболической инженерии. Adv. Biochem. Англ. Biotechnol. 148: 207-239. DOI: 10.1007 / 10_2014_301
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пхукан, У. Дж., Джина, Г. С., и Шукла, Р. К. (2016). Факторы транскрипции WRKY: молекулярная регуляция и стрессовые реакции у растений. Фронт. Plant Sci. 7: 760. DOI: 10.3389 / fpls.2016.00760.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Расул, С.и Мохамед Р. (2016). «Понимание образования агарового дерева и его проблем», в Agarwood: Science Behind the Fragrance , ed. Р. Мохамед. (Берлин: Springer), 39–56. DOI: 10.1007 / 978-981-10-0833-7_3
CrossRef Полный текст | Google Scholar
Рейс, Дж., Гаспар, А., Милхазес, Н., и Борхес, Ф. (2017). Хромон как основа для открытия новых лекарств: последние достижения. J. Med. Chem. 60, 7941–7957. DOI: 10.1021 / acs.jmedchem.6b01720.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ромер, М.(1999). Открытие независимого от мевалоната пути биосинтеза изопреноидов у бактерий, водорослей и высших растений. Нац. Prod. Rep. 16, 565–574. DOI: 10.1039 / a709175c
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сангаресвари Нагаджоти, М., Тангамуту Партхибан, К., Умеш Канна, С., Картиба, Л., и Сараванакумар, Д. (2016). Грибковые микробы, связанные с образованием агарового дерева. г. J. Plant Sci. 7, 1445–1452. DOI: 10.4236 / ajps.2016.710138
CrossRef Полный текст | Google Scholar
Schmiesing, A., Emonet, A., Gouhier-Darimont, C., and Reymond, P. (2016). Факторы транскрипции MYC арабидопсиса являются мишенью гормонального перекрестного взаимодействия салициловая кислота / жасмоновая кислота в ответ на экстракт яйца Pieris brassicae. Plant Physiol. 170: 2432. DOI: 10.1104 / стр. 16.00031
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шао, Х., Мэй, W. -L., Dong, W. -H., Gai, C. -J., Li, W., Zhu, G. -P., Et al. (2016). Производные 2- (2-фенилэтил) хромона древесного агара, происходящие из Gyrinops salicifolia . Молекулы 21: E1313. DOI: 10,3390 / молекулы21101313.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шимада Ю., Томинага Т. и Киёсава С. (1986). Исследования агалового дерева (Джинко). IV: корреляция между классификацией древесины агалового дерева на рынке и производными хромона. Yakugaku Zasshi J. Pharm. Soc. Jpn. 106, 391–397. DOI: 10.1248 / yakushi1947.106.5_391
CrossRef Полный текст | Google Scholar
Шимада Ю., Томинага Т., Кониси Т. и Киёсава С. (1982). Исследования на агаровом дереве (Джинко). I. Строение производных 2- (2-фенилэтил) хромона. Chem. Pharm. Бык. 30, 3791–3795. DOI: 10.1248 / cpb.30.3791.
CrossRef Полный текст | Google Scholar
Сибуриан Р. Х., Сирегар У. Дж., Сирегар И. З., Сантосо Э. (2015). Идентификация морфологических признаков Aquilaria microcarpa при взаимодействии с Fusarium solani . Внутр. J. Sci. Базовое приложение Res. 20: 119–128.
Google Scholar
Сингх Б. и Шарма Р. А. (2015). Терпены растений: защитные реакции, филогенетический анализ, регуляция и клиническое применение. 3 Biotech. 5, 129–151. DOI: 10.1007 / s13205-014-0220-2.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Синха, А. К., Джагги, М., Рагурам, Б., и Тутеджа, Н. (2011). Передача сигналов митоген-активируемой протеинкиназы у растений в условиях абиотического стресса. Завод Сигнал. Behav. 6, 196–203. DOI: 10.4161 / psb.6.2.14701.
CrossRef Полный текст | Google Scholar
Субасингхе, С.М.С.У.П., Хеттиараччи, Д.С. (2015). Характеристика смолы типа агарового дерева Gyrinops walla Gaertn, произрастающей в отдельных популяциях в Шри-Ланке. Ind. Crops Prod. 69, 76–79. DOI: 10.1016 / j.indcrop.2015.01.060.
CrossRef Полный текст | Google Scholar
Subasinghe, U., и Hettiarachchi, D.(2013). Производство смолы Agarwood и качество смолы Gyrinops walla Gaertn. Внутр. J. Agr. Sci. 3, 357–362.
Google Scholar
Тауфик, Х.А., Эвис, Э. Ф., и Эль-Хамули, В. С. (2014). Синтез хромонов и их применение за последние десять лет. Внутр. J. Res. Pharm. Chem. 4, 1046–1085.
Google Scholar
Тутея, Н., и Махаджан, С. (2007). Сеть сигнализации кальция в растениях: обзор. Завод Сигнал.Behav. 2, 79–85. DOI: 10.4161 / psb.2.2.4176
CrossRef Полный текст | Google Scholar
Ван Тхань, Л., Ван До, Т., Сон, Н. Х., Сато, Т., и Козан, О. (2015). Влияние биологической, химической и механической обработки на содержание сесквитерпена в стеблях посаженных деревьев Aquilaria crassna . Агролесоводство. Syst. 89, 973–981. DOI: 10.1007 / s10457-015-9829-3.
CrossRef Полный текст | Google Scholar
Ван, С. Л., Хван, Т. Л., Чунг, М.I., Sung, P.J., Shu, C.W., Cheng, M.J., et al. (2015). Новые флавоны, производное 2- (2-фенилэтил) -4H-хромен-4-она и противовоспалительные компоненты коры ствола Aquilaria sinensis . Molecules 20, 20912–20925. DOI: 10,3390 / молекулы201119736.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ван Т., Ли, Л. Ф., Чжан, К., Чжан, В. Ю. и Пей, Ю. Х. (2001). Новые 2- (2-фенилэтил) хромоны из Bothriochloa ischaemum . J. Asian Nat. Prod. Res. 3, 145–149. DOI: 10.1080 / 10286020108041382.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ван Х., Гао Б., Лю Х., Дун Х., Чжан З., Фань Х. и др. (2016). Солевой стресс индуцирует продукцию 2- (2-фенилэтил) хромонов и регулирует новые классы чувствительных генов, участвующих в передаче сигнала в каллусах Aquilaria sinensis . BMC Plant Biol. 16: 119. DOI: 10.1186 / s12870-016-0803-7.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wu, B., Lee, J., Lim, C., Dong Jia, S., Won Kwon, S., Seo Hwang, G., et al. (2012a). Сесквитерпеноиды и производные 2- (2-фенилэтил) -4H-хромен-4-она (= 2- (2-фенилэтил) -4H-1-бензопиран-4-он) из агарового дерева Aquilaria malaccensis . Helv. Чим. Acta 95, 636–642. DOI: 10.1002 / hlca.201100409
CrossRef Полный текст | Google Scholar
Ву, Б., Вон Квон, С., Со Хван, Г., и Хилл Парк, Дж. (2012b). Восемь новых производных 2- (2-фенилэтил) хромона (= 2- (2-фенилэтил) -4H-1-бензопиран-4-он) из агарового дерева Aquilaria malaccensis . Helv. Чим. Acta 95, 1657–1665. DOI: 10.1002 / hlca.201200069.
CrossRef Полный текст | Google Scholar
Ву, З. К., Лю, С., Ли, Дж. Ф., Ли, М. К., Ду, Х. Ф., Ци, Л. К. и др. (2017). Анализ экспрессии генов и качества древесины агара с использованием Agar-bit в Aquilaria sinensis . Дж.Троп. Для. Sci. 29, 380–388. DOI: 10.26525 / jtfs2017.29.3.380388
CrossRef Полный текст | Google Scholar
Сюй, К., Лю, Р., Чжан, К., Чен, X., Цянь, Ю. и Фанг, В. (2017). Диверсификация эволюционно консервативных каскадов MAPK коррелирует с эволюцией видов грибов и развитием образа жизни. Genome Biol. Evol. 9, 311–322. DOI: 10.1093 / GBE / evw051.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сюй, Ю.-H., Liao, Y. -C., Zhang, Z., Liu, J., Sun, P. -W., Gao, Z. -H., Et al. (2016). Жасмоновая кислота является важным преобразователем сигнала в индуцированном тепловым шоком образовании сесквитерпена у Aquilaria sinensis . Sci. Реп. 6: 21843. DOI: 10.1038 / srep21843
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Xu, Y., Zhang, Z., Wang, M., Wei, J., Chen, H., Gao, Z., et al. (2013). Идентификация генов, связанных с образованием агарового дерева: анализ транскриптома здоровых и раненых тканей Aquilaria sinensis . BMC Genomics 14: 227. DOI: 10.1186 / 1471-2164-14-227.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Xu, Y.H., Liao, Y. C., Lv, F. F., Zhang, Z., Sun, P. W., Gao, Z. H., et al. (2017). Фактор транскрипции AsMYC2 контролирует чувствительную к жасмонату экспрессию ASS1, регулирующую биосинтез сесквитерпена, у Aquilaria sinensis (Lour.) Gilg. Physiol растительных клеток. 58, 1924–1933. DOI: 10.1093 / pcp / pcx122.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сюй, Ю.Х., Ван, Дж. У., Ван, С., Ван, Дж. Й. и Чен, X. Y. (2004). Характеристика GaWRKY1, фактора транскрипции хлопка, который регулирует ген сесквитерпенсинтазы (+) — дельта-кадиненсинтаза-A. Plant Physiol. 135, 507–515. DOI: 10.1104 / стр.104.038612.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ян Д. Л., Ван, Х., Го, З. К., Донг, В. Х., Мэй, В. Л. и Дай, Х. Ф. (2014a). Новое производное 2- (2-фенилэтил) хромона в китайском агаровом дереве ‘Qi-Nan’ из Aquilaria sinensis . J. Asian Nat. Prod. Res. 16, 770–776. DOI: 10.1080 / 10286020.2014.896342.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ян, Д. -Л., Ван, Х., Го, З. -К., Ли, В., Мэй, В. -Л. И Дай, Х. -Ф. (2014b). Ароматный агарофуран и сесквитерпены эремофилана в древесине агара «Qi-Nan» из Aquilaria sinensis . Phytochem. Lett. 8, 121–125. DOI: 10.1016 / j.phytol.2014.03.003.
CrossRef Полный текст | Google Scholar
Ян, Л., Qiao, L., Xie, D., Yuan, Y., Chen, N., Dai, J., et al. (2012). 2- (2-фенилэтил) хромоны из китайского иглвуда. Phytochem. 76, 92–97. DOI: 10.1016 / j.phytochem.2011.11.017.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ян, М., Фу, Х., Лян, Ю., Хуанг, Х., Чжао, Б., Се, К. и др. (2014c). Модифицированные устройства для переливания, индуктор и процедура для индуцирования агарового дерева методом инфузии. J. Chem. Pharm. Res. 6, 2566–2571.
Google Scholar
Ян, X., Вэй, Дж. Х., Лю, Дж., И Сюй, Ю. Х. (2013). Клонирование и анализ экспрессии фарнезилпирофосфатсинтазы из Aquilaria sinensis . Чжунго Чжун Яо За Чжи. 38, 3251–3255.
PubMed Аннотация | Google Scholar
Ye, W., He, X., Wu, H., Wang, L., Zhang, W., Fan, Y., et al. (2018). Идентификация и характеристика новой сесквитерпен-синтазы из Aquilaria sinensis : важный ген для образования агарового дерева. Внутр.J. Biol. Макромол. 108, 884–892. DOI: 10.1016 / j.ijbiomac.2017.10.183.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ye, W., Wu, H., He, X., Wang, L., Zhang, W., Li, H., et al. (2016). Секвенирование транскриптома химически индуцированного Aquilaria sinensis для идентификации генов, связанных с образованием агарового дерева. PLoS One 11: e0155505. DOI: 10.1371 / journal.pone.0155505.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Йосватана, Н.(2013). Экстракция масла агарового дерева ( Aquilaria crassna ) с использованием сверхкритической экстракции диоксидом углерода и предварительной обработки ферментами при гидродистилляции.